| Скачать .docx | Скачать .pdf |
Реферат: Принципы биохимического исследования
Реферат
Принципы биохимического исследования
Введение
Основная задача биохимии — объяснить, как функционируют живые системы с точки зрения процессов, протекающих в клетках. Все клетки в организме находятся в состоянии динамической активности и подвергаются действию внутренних и внешних факторов, которые в свою очередь также постоянно изменяются. В процессе жизнедеятельности любая отдельно взятая клетка взаимодействует с другими клетками, находящимися как в непосредственной близости от нее (межклеточные взаимодействия), так и на некотором расстоянии (гормональные эффекты). Функционирование органеллы внутри клетки также в значительной степени зависит от активности других органелл и окружающей цитоплазмы. Ясно поэтому, что нельзя достаточно полно изучить живую клетку, если делать это в отрыве от целого организма.
Интенсивные исследования, расширяющие и углубляющие наши знания о многочисленных процессах, протекающих в живом организме, ведутся в области фармакологии, микробиологии, патологии и других наук. Биохимия изучает эти процессы главным образом на уровне клетки и клеточных структур, однако полученные результаты должны рассматриваться также на уровне органов, тканей, всего организма и даже во взаимосвязи организма с окружающей средой.
Изучение любой последовательности взаимосвязанных процессов, протекающих в какой-либо биологической системе, начинают, как правило, с изучения ее компонентов. Для этого компоненты обычно выделяют, всесторонне их исследуют и пытаются понять, как они функционируют в составе организма.
Процессы, характерные для целой клетки, протекают в отдельных клеточных частицах и органеллах, которые для анализа выделяют из клетки с помощью фракционирования. Этот процесс обычно состоит из двух этапов (разд. 1.10.3): сначала клетки разрушают, а затем из образовавшейся суспензии методом центрифугирования (гл. 2) выделяют нужные частицы и органеллы. Дальнейшее разделение индивидуальных компонентов клеточных частиц и органелл и изучение их свойств проводят с помощью центрифугирования (гл. 2), хроматографии (гл. 3) или электрофореза (гл. 4). Для определения состава, механизма действия и функций клеточных компонентов пользуются сложными количественными и качественными аналитическими методами. На атомном и молекулярном уровнях применяют целый ряд спектральных методов (гл. 5); механизм действия клеточных частиц и внутриклеточные взаимодействия изучают, используя одновременно несколько аналитических методов, таких, как спектроскопия (гл. 5) и радиоизотопные методы (гл. 6), потенциометрия, полярография (гл. 7) и манометрия (гл. 8).
При фракционировании нормальная активность клетки может в значительной степени нарушаться. Чтобы свести последствия фракционирования до минимума и приблизить условия к естественным, применяют особые приемы (разд. 1.10.1). Однако все побочные явления, возникающие в ходе фракционирования, устранить невозможно, поэтому полученные результаты следует трактовать весьма осторожно, особенно если речь идет о целой клетке, органе или организме.
1. рН среды и буферные растворы
1.1 Влияние рН на биологические процессы
Организмы и клетки, как правило, весьма устойчивы даже к значительным изменениям рН окружающей среды. Внутриклеточные процессы, наоборот, обладают высокой чувствительностью к рН и протекают в среде, рН которой строго регулируется (правда, некоторые колебания рН могут наблюдаться и внутри клетки, например у поверхности мембраны). Большинство внутриклеточных процессов протекает при нейтральных значениях рН, когда их скорость максимальна. Гидролазы лизосом, однако, обладают максимальной активностью при рН 5,0. Желудочный сок млекопитающих имеет весьма необычную величину рН — около 1; именно при этом рН активность фермента пепсина, начинающего переваривание белков пищи в желудке, максимальна.
В биологических системах постоянная величина рН поддерживается с помощью эффективных буферных систем, которые по своей химической природе таковы, что они препятствуют изменениям рН, возникающим в ходе метаболического образования кислот (например, молочной кислоты) и оснований (например, аммиака). Большинство буферных систем, содержащихся в клеточных жидкостях, включают фосфаты, бикарбонат, аминокислоты и белки.
Чувствительность биологических процессов к рН обусловлена целым рядом причин. Ионы водорода могут выступать в качестве катализатора ряда процессов, быть реагентом или продуктом реакции. Кроме того, при изменении рН может измениться проницаемость клеточной мембраны, а следовательно, и распределение веществ или ионов по обе ее стороны. Подобно другим биологическим структурам, мембраны содержат способные к ионизации группы, и в зависимости от степени их ионизации меняется конформация, а значит и биологическая активность молекул, в которые эти группы входят. Это прежде всего касается белков, а следовательно, ферментов. В некоторых белках небольшое изменение рН окружающей среды вызывает проявление биологической активности. На примере гемоглобина, основной функцией которого является перенос кислорода от легких к тканям, можно видеть, что при активном тканевом дыхании незначительное понижение рН в тканях в результате образования углекислоты и ионов водорода облегчает высвобождение кислорода. Процесс высвобождения кислорода сопровождается связыванием протонов гемоглобином, что увеличивает буферную емкость системы.
При изучении метаболических процессов in vitro возникает необходимость в применении «нефизиологических» буферных растворов: направленное изменение рН может значительно облегчить изучение таких типов молекул, как аминокислоты, белки и нуклеиновые кислоты с помощью электрофореза и ионообменной хроматографии.
1.2 Буферные растворы для биологических исследований
Буферным называется такой раствор, который препятствует изменению концентрации ионов водорода при добавлении к нему кислоты или щелочи. Такое действие раствора называется буферным. Величину буферного действия характеризуют буферной емкостью р, равной количеству сильного основания, которое необходимо добавить для изменения рН раствора на одну единицу.
![]()
где d(pH) — изменение рН раствора при добавлении основания.
Обычно пользуются буферными растворами, состоящими из смеси слабой кислоты или основания и соли этой кислоты, например смеси уксусной кислоты и ацетата натрия (по номенклатуре Брёнстеда и Лоури буферный раствор представляет собой смесь слабой кислоты и сопряженного с ней основания).
При добавлении к раствору слабой кислоты (НА) и ее соли (А~) ионов водорода последние нейтрализуются анионами соли, которые действуют в данном случае как слабое основание; гидроксильные ионы, наоборот, нейтрализуются кислотой. Отсюда следует, что буферная емкость раствора, составленного из данной кислоты и сопряженного с ней основания, максимальна в том случае, когда их концентрации равны, т. е. рН = рКа кислоты. Буферная емкость зависит также от общей концентрации раствора и отношения соль— кислота: чем выше концентрация раствора, тем больше его буферная емкость. Концентрация кислоты и соли в буферных растворах обычно бывает порядка 0,05—0,20 М, а достаточной буферной емкостью растворы обладают в области значений
рН = рКа ± 1
Буферы, применяемые для биологических исследований, должны удовлетворять ряду требований:
1. Обладать достаточной буферной емкостью в требуемом диапазоне значений рН.
2. Обладать высокой степенью чистоты.
3. Хорошо растворяться в воде и не проникать через биологические мембраны.
4. Обладать устойчивостью к действию ферментов и гидролизу.
5. рН буферных растворов должен как можно меньше зависеть от их концентрации, температуры и ионного или солевого состава среды.
6. Не оказывать токсического или ингибирующего действия.
7. Комплексы буфера с катионами должны быть растворимыми.
8. Не поглощать свет в видимой или ультрафиолетовой областях спектра.
К сожалению, этим требованиям удовлетворяют далеко не все буферные растворы. Так, фосфаты обладают способностью осаждать поливалентные катионы и во многих системах выступают в качестве метаболитов или ингибиторов; трис-буфер иногда оказывает токсическое или ингибирующее действие. До недавнего времени насчитывалось всего несколько буферов, рН которых лежит в важной для биохимии области 6,0—8,0 и которые удовлетворяют перечисленным выше требованиям. В последние годы, однако, появился целый ряд так называемых цвиттерионных буферов типа ГЭПЭС и ПИГТЭС. Некоторые наиболее распространенные буферы приведены в табл. 1.1. Для получения буферных растворов, применимых в широком диапазоне значений рН, используются смеси разных буферов. Например, буферы Мак-Ильвейна имеют рН с областью значений от 2,2 до 8,0 и приготавливаются из лимонной кислоты и двузамещенного фосфорнокислого натрия.
Наиболее важной группой физиологических буферов являются белки. Благодаря большому количеству содержащихся в боковых цепях аминокислот щелочных и слабокислых групп белки имеют очень высокую буферную емкость. Буферная емкость крови в основном определяется гемоглобином.
1.3 Зависимость ионизации аминокислот и белков от рН
Аминокислоты и белки — это наиболее важные в биологическом отношении соединения, поэтому необходимо знать, в какой степени изменение рН влияет на их физические свойства. За исключением пролина, химические формулы всех аминокислот, из которых синтезируются белки, можно записать в общем ' виде как RCH(NH2 )COOH. рН аминогруппы лежит в области 9,0—10,5, а карбоксильной группы — между 1,7 и 2,4.
Степень ионизации аминокислот в водных растворах зависит от рН и определяется уравнением Гендерсона—Хассельбальха.
![]()
 |
Таким образом, при низких значениях рН аминокислота находится в катионной форме, а при высоких — в анионной. При некотором промежуточном значении рН аминокислота оказывается незаряженной и называется цвиттерионом. Было установлено, что в кристаллическом состоянии или после растворения в чистой воде такие аминокислоты существуют главным образом в виде цвиттерионов, что придает им свойства ионных соединений, а именно высокую точку плавления и кипения, хорошую растворимость в воде и плохую растворимость в таких органических растворителях, как эфир и хлороформ. Величина рН, при которой в водном растворе преобладает цвиттерион, называется изоионной точкой: число отрицательных зарядов, образующихся на молекуле в результате отщепления протонов, равно числу положительных зарядов, образующихся благодаря присоединению протонов. Для аминокислот эта величина приблизительно соответствует изоэлектрической точке (pi) — молекула не несет суммарного заряда и таким образом оказывается электрофоретически неподвижной. Численное значение рН для этого случая зависит от того, насколько сильной является кислота, и определяется следующим уравнением:
Для глицина величины рКа, Н рКаг равны 2, 3 и 9,6 соответственно; следовательно, изоионная точка равна 6,0. При более низких значениях рН в растворе содержатся и цвиттерион, и катион, а соотношение между ними определяется уравнением Гендерсона — Хассельбальха; при более высоких рН наряду с цвиттерионом в растворе находится анион.
Для так называемых «кислых» аминокислот, например, аспарагиновой кислоты; ионизация носит несколько иной характер. Это обусловлено наличием у них второй карбоксильной группы.
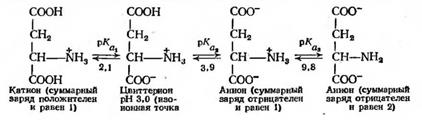
В этом случае рН, при котором в водном растворе преобладает цвиттерион, определяется величинами рСа , и рКаз
Для лизина, который является «основной» аминокислотой, изоионная точка определяется величинами рКа2 и рКа3 - Ионизация происходит следующим образом:
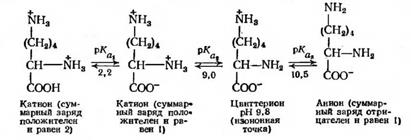
Вместо второй амино- или карбоксильной группы боковая цепь аминокислоты иногда содержит другую химическую группу, которая при определенном значении рН также ионизуется. К таким группам относятся фенольная (тирозин), гуанидиновая (аргинин), имидазольная (гистидин) и сульфгидрильная группа (цистеин). Ясно, что степень ионизации различных основных групп аминокислот при одном и том же рН будет различной. Более того, небольшие различия могут наблюдаться даже у одной и той же группы. Эти различия используют при электрофоретическом и ионообменном разделении смесей аминокислот, имеющихся, например, в белковом гидролизате.
Ионизация молекул белка качественно напоминает ионизацию аминокислот, но в количественном отношении отличается от нее благодаря наличию большого числа способных к ионизации групп. Белки образуются путем конденсации а-аминогруппы одной аминокислоты с о-карбоксильной группой соседней аминокислоты, поэтому, за исключением двух концевых аминокислот, все о амино- и карбоксильные группы участвуют в образовании пептидных связей и в белке не ионизуются. Однако в боковых цепях присутствуют сотни амино- и карбоксильных групп, которые могут легко ионизоваться. Электростатическое притяжение, возникающее между некоторыми из этих групп, способствует стабилизации третичной структуры белковой молекулы, при этом молекулы белков часто бывают свернуты таким образом, что большинство способных к ионизации групп оказываются расположенными на поверхности молекулы, где они могут вступать во взаимодействие с окружающей средой. Естественно, что относительное число положительно и отрицательно заряженных групп в молекуле белка определяет те или иные ее физические свойства. У гистонов преобладают катионные группы, в то время как в других белках количество анионных и катионных групп либо одинаково, либо, напротив, преобладают анионные группы.
В отличие от аминокислот у белков изоионная точка обычно не совпадает с изоэлектрической. По определению изоионная точка — это такая величина рН, при которой молекула белка содержит равное число положительно и отрицательно заряженных групп, образовавшихся в результате связывания основных групп с протонами и соответствующей диссоциации кислотных групп. Изоэлектрическая точка — это рН, при котором белок электрофоретически неподвижен. При ее определении белок растворяют в буферном растворе. В этом растворе всегда содержатся низкомолекулярные анионы и катионы, способные связываться с многочисленными заряженными группами белка. В таких условиях наблюдаемое в изоэлектрической точке равновесие зарядов частично обусловлено их компенсацией связавшимися с молекулой белка анионами и катионами.
Молекулы белка всегда изучают в буферных растворах; при этом важно установить изоэлектрическую точку, поскольку, на пример, именно при этом значении рН создаются наиболее благоприятные условия для взаимодействия между противоположно заряженными группами соседних молекул, что ведет к их последующей агрегации и быстрому осаждению.
Агрегацию, а следовательно, и осаждение белковых молекул можно вызвать добавлением к раствору белка солей, например сульфата аммония. Молекулы белка и неорганические ионы при гидратации конкурируют за молекулы воды, и по достижении определенной концентрации соли взаимодействия белок — белок начинают преобладать над взаимодействиями белок — вода, что приводит к агрегации, а затем и осаждению белка. Этот метод широко применяется на начальных стадиях очистки белков. Различия в изоэлектрической точке разных белковых молекул используются при разделении белков с помощью электрофореза.
2. Подходы к биохимическому исследованию
Как мы уже говорили, активность клетки зависит от взаимодействия ее как с непосредственным окружением, так и с клетками, значительно удаленными от нее; кроме того, играет роль также и то, какие методы применяются для ее изучения. Поэтому чтобы получить наиболее полное представление о состоянии клеток внутри органов и тканей, опыты лучше всего проводить на интактных изолированных органах и тканях. Преимущество такого изучения заключается в том, что при этом удается избежать нежелательных эффектов, обусловленных изменением непосредственного тканевого окружения, как это имеет место, например, при фракционировании клеток. Исследование на уровне целых организмов не только не нарушает целостности тканей, но позволяет изучать отдельные органы и ткани, оставляя без изменения снабжение их питательными веществами, нервную и гормональную регуляцию и т. д. Таким образом, чтобы получить полную картину клеточного обмена, необходимо провести исследование на целом организме, изолированном органе, на уровне клетки, клеточной органеллы, на молекулярном и атомном уровнях.
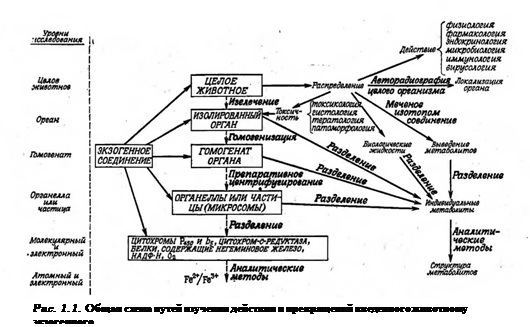
На рис. 1.1 показаны пути биохимических подходов к изучению возможных превращений какого-либо экзогенного соединения (ксенобиотика) в организме животного или его действия на этот организм. Существуют два подхода, которые взаимно дополняют друг друга.
Первый из них заключается в том, что животному вводят соединение, а затем выделяют и идентифицируют все продукты, выводимые из организма. В других случаях спустя некоторое время после введения соединения животное умерщвляют, выделяют и фракционируют нужный орган или ткань, а затем изучают судьбу введенного соединения или исследуют его действие на интересующую субклеточную систему.
Второй подход состоит во введении соединения на одном из указанных на рис. 1.1 уровнях (т. е. в соответствующий изолированный орган, гомогенат, органеллу или субклеточную систему) и изучении действия этого соединения или путей его превращения на данном или более низких уровнях.
Действие какого-либо соединения можно исследовать на нескольких клеточных частицах или органеллах, каждую из которых необходимо для этого выделить из системы и подвергнуть анализу.
Зачастую исследуемое соединение представляет собой природный метаболит или входит в состав самого организма, а иногда является изотопом составного компонента организма. С другой стороны, не всегда действие какого-либо экзогенного соединения на функционирование организма представляет интерес для биохимика. В любом случае изучение можно проводить на любом из вышеуказанных уровней организации, а получаемые на каждом уровне результаты помогают составить более полное представление о процессах, протекающих в организме.
3. Исследования на уровне целого организма
Биохимические эксперименты на животных могут быть предприняты с различными целями. Применяя в течение длительного времени специальную диету, лишенную определенных витаминов или микроэлементов, и одновременно регистрируя возникающие при этом физиологические и клинические изменения, можно исследовать метаболическую роль данного витамина или микроэлемента. Вместе с тем, вводя животному какое-либо экзогенное соединение, можно изучать как влияние этого соединения на организм животного (фармакологическая или патологическая ответная реакция), так и влияние организма на введенное соединение, т. е. его превращения и выведение. В последнее время, главным образом благодаря созданию комитета Данлопа (ныне Комитет по безопасности применения лекарственных средств) после известной трагедии с талидомидом, все большее внимание стали уделять вопросам о том, каким образом различные лекарственные вещества, пищевые добавки и красители, а также такие соединения, как ДДТ, накапливаются в пище и метаболизируются, где они локализуются, какое разрушающее действие оказывают на органы и ткани и какие тератогенные и канцерогенные побочные эффекты могут вызывать.
Единственным способом исследовать пути превращения введенных соединений у человека является определение содержания этих соединений и их метаболитов в крови, моче, фекалиях, желчи, выдыхаемом воздухе, поте и слюне. О поражении органа судят по изменению содержания в сыворотке крови таких ферментов, как аспартат—аминотрансфераза и лактатдегидрогеназа. После введения соединений лабораторных животных умерщвляют, а затем исследуют изолированные органы.
Результаты, получаемые при анализе содержания каких-либо соединений и их метаболитов в биологических жидкостях и экскрементах живых организмов, в том числе и организма человека, зависят от многих взаимосвязанных факторов. К этим факторам относятся:
Способ введения соединения (перорально или путем внутривенной, внутримышечной, подкожной или внутрибрюшинной инъекции).
Скорость всасывания соединения в кровоток; если соединение вводят не внутривенно, то оно должно проникнуть (обычно путем пассивной диффузии) по крайней мере через одну мембрану (если соединение представляет собой слабый электролит, то оно должно пройти через мембрану в неионизованной форме).
Степень связывания соединения с сывороточным альбумином.
Кровоснабжение тканей.
Способность данного соединения проходить через мембраны и распределяться в межклеточных и внутриклеточных жидкостях.
Накопление соединения в жировой ткани и характер его связывания с нуклеиновыми кислотами и меланином.
Скорость метаболизма соединения (главным образом ферментной системой микросом печени).
Скорость выведения (с мочой и желчью).
Совокупность вышеперечисленных факторов создает характер.
Для данного соединения «фармако-кинетическую» картину. Помимо всего перечисленного, имеют значение также возраст, пол, режим питания, физические нагрузки, генетическое строение животного, а также время дня, когда животному было введено соединение (ввиду наличия у животных циркадного ритма). Оказалось даже, что превращение введенного соединения у животных, содержащихся в клетках с мягкими опилками, происходит быстрее, чем у животных, помещенных в клетки с жесткими опилками. Именно благодаря огромному многообразию переменных эксперименты всегда следует проводить не на одном животном, а на целых группах. Исследования нужно проводить на животных чистых линий, а получаемые экспериментальные данные подвергать тщательной статистической обработке. Результаты анализа фармакологического действия введенных экспериментальным животным соединений необходимо сопоставлять с данными, получаемыми в группе контрольных животных, которым было введено плацебо, т. е. совершенно нейтральное вещество, например лактоза.
Изучение метаболизма ксенобиотиков проводят на самых разных > лабораторных животных — мышах, крысах, морских свинках (из-за их малой стоимости и простоты ухода за ними); наряду с ними для опытов используют также кроликов, кошек, собак и обезьян. Одной из основных проблем, стоящих перед биохимиками и фармакологами, является экстраполяция результатов, полученных на лабораторных животных, к человеку. Многочисленные сравнительные исследования показали, что метаболические процессы у человека и всех исследованных видов животных существенно различаются. Именно по этой причине новые лекарственные препараты, предназначенные для введения человеку, предварительно проходят испытание не на одном животном, а на целом ряде различных лабораторных животных. Только после тщательного сравнительного анализа полученных результатов клинические испытания можно проводить на человеке.
При изучении метаболизма ксенобиотиков удобнее всего вводить их в виде изотопных меток; как правило, вводят препараты, меченные по 14 0 С. Экскрецию у мелких лабораторных животных целесообразно изучать с помощью специальной метаболической стеклянной камеры, в которую животное помещают на все время опыта (рис. 1.2). Благодаря особой конструкции камеры моча и фекалии собираются отдельно, а выдыхаемый углекислый газ поступает в специальную ловушку. При изучении меченных по углероду соединений в клетку подается воздух, не содержащий углекислого газа, а выдыхаемый углекислый газ поглощается раствором гидроокиси натрия; анализ выдыхаемого углекислого газа, меченного по углероду (14 С02 ), позволяет установить степень распада изучаемого соединения. Выделяемые с мочой и фекалиями вещества и их метаболиты связаны, как правило, с такими соединениями, как Р-глюкуроновая кислота, глицин, глутатион и сульфат, что повышает растворимость метаболитов в воде, а следовательно, и степень их экскреции. Поэтому перед экстракцией метаболитов органическими растворителями (эфиром или хлороформом) пробы экскрементов нужно гидролизовать, предварительно исследовав их соответствующими аналитическими методами.
Одним из способов изучения путей превращения введенного соединения in vivo является перфузия органов (например, почек или печени), при которой соединение вводят с помощью тонкой полой иглы в артерию, несущую кровь к данному органу, а затем анализируют пробы крови, взятые из соответствующей вены животного.
В Англии существуют инструкции, ограничивающие применение живых позвоночных животных для исследовательских целей; контроль за выполнением этих инструкций осуществляется специальными комиссиями Внутреннего департамента (Министерства внутренних дел Великобритании). Эксперименты на живых позвоночных нельзя проводить без особого разрешения министерства, которое должно быть подписано профессором физиологии (или специалистом смежного профиля) и президентом Королевского общества или его заместителем.
Для изучения распределения введенного соединения животное спустя некоторое время после инъекции умерщвляют с помощью анестезий, декапитации или цервикальной дислокации. Затем проводят исследования либо трупа животного, либо отдельных органов, которые для этой цели изолируют, выявляют морфологические изменения, происшедшие в них, изучают их составные части.
Для получения наглядного представления о распределении введенных мелким лабораторным животным радиоактивных соединений успешно применяется метод авторадиографии препаратов целого организма (разд. 6.2.4). Этот метод позволяет получить данные о распределении и относительном содержании введенного соединения в тканях животного, скорости его выведения и способности проникать сквозь биологические мембраны. Через определенный промежуток времени после введения соединения животное умерщвляют с помощью анестезии и быстро замораживают смесью ацетона с твердым С02 при температуре —78° С или с помощью жидкого азота. Замороженное животное помещают при низкой температуре в водный раствор смолы (аравийская камедь), а после застывания смолы рассекают на соответствующем уровне резцом, прикрепленным к электродрели, или делают секционные срезы с помощью микротома со специальным лезвием из карбида вольфрама. Полученный срез прикладывают к рентгеновской пленке и оставляют на 1—2 нед при низкой температуре, после чего пленку проявляют. Сопоставляя полученную авторадиограмму с цветным снимком среза, изучают распределение и локализацию введенного животному изотопа в различных его органах и тканях. Типичная авторадиограмма изображена на рис. 1.3.
4. Изотонические солевые растворы
При проведении экспериментов на органах животных, срезах растительных и животных тканей, гомогенатах и клеточных органеллах необходимо, чтобы среда для суспендирования имела не только определенный рН, но и заданный ионный состав. Среда должна быть изотонической, т. е. осмотическое давление в ней должно совпадать с осмотическим давлением внутри клетки или клеточной органеллы, чтобы их метаболическая целостность не нарушалась. Кроме того, если, например, изучают рост клеток, среда для суспендирования должна содержать все необходимые основные питательные вещества. Это требование нужно особенно тщательно выполнять при изучении культур клеток и тканей, особенно культур животных клеток.
Существует целый ряд физиологических солевых растворов, многие из которых являются разновидностями одного из первых — раствора Рингера. К ним относятся растворы Тайрода, Янга, Лока, Менга и Да Жалона. Наиболее часто применяются фосфатный и бикарбонатный растворы Кребса—Рингера. По своему ионному составу бикарбонатный солевой раствор близок к сыворотке крови млекопитающих. Фосфатный раствор Кребса—Рингера не является физиологическим, но может с успехом применяться для изучения срезов и гомогенатов тканей в тех случаях, когда поглощение кислорода измеряют манометрическими методами (разд. 8.6.2). Большинство этих солевых растворов содержат в различных количествах NaCl, КС1, MgS04 , СаС12 , NaHCOs и КН2 Р04 ; некоторые растворы насыщены смесью газов — кислорода и углекислого газа. Кроме того, в их состав могут входить такие соединения, как глюкоза, пируват, фумарат и оксалоацетат. Для исследования культур животных тканей используются растворы Хэнкса и Гей—Эрля. Выбор того или иного раствора вначале является произвольным и не основывается на объективных данных, поэтому перед его применением необходимо убедиться, что состав его оптимален. И, наконец, при проведении физиологических экспериментов нужно следить за тем, чтобы эффекты, наблюдаемые при изменении рН среды, были вызваны изменениями в концентрации ионов водорода, а не какими-либо другими изменениями буферного раствора.
5. Перфузия изолированных органов
Сущность этого метода заключается в том, что изучаемый орган (печень, почку или сердце) изолируют из организма животного и помещают в специальный термостатируемый прибор. Затем к перфузионной жидкости, которая обычно вводится в орган через артерию, добавляют исследуемое соединение и анализируют жидкость, вытекающую из органа через вену, что позволяет проследить за превращениями введенного соединения. Перфузионную жидкость можно пропускать через орган однократно или несколько раз, самотеком или с помощью небольшого насоса. Насос применяют тогда, когда перфузионную жидкость пропускают через орган многократно. В отдельных случаях жидкость прогоняют не при постоянном давлении, а импульсами, что позволяет приблизить условия опыта к ситуации in vivo и имитирует процесс перекачивания крови сердцем.
Для проведения перфузии не обязательно полностью изолировать орган; ее можно проводить и на органе вскрытого анестезированного животного. При этом удается сохранить интактными нервные волокна и часть сосудистой системы.
Действие какого-либо соединения на ткань или орган (например, гистамина на мышцу) можно изучать по механической ответной реакции изолированной ткани на данное соединение. Исследуемое соединение добавляют к омывающей ткань жидкости; в ответ на это ткань, закрепленная с одного конца, а другим концом связанная с пером самописца, начинает двигаться, и любое движение ткани постоянно регистрируется на самописце. Данный метод позволяет изучать ответную реакцию изолированных органов на введение очень небольших (порядка нескольких нанограмм) количеств активных соединений. Основным недостатком метода перфузии изолированных органов является отсутствие гормонального и нервного контроля; поэтому при экстраполяции всех полученных результатов к ситуации in vivo следует соблюдать осторожность.
6. Приготовление срезов органов и тканей
Срезы тканей желательно делать как можно тоньше, чтобы обеспечить свободный доступ кислорода в самые глубокорасположенные слои срезов и полное выведение продуктов распада за счет диффузии. Этим требованиям удовлетворяют срезы толщиной от 0,5 до 5 мм; кроме того, в этом случае соотношение между разрушенными и интактными клетками остается достаточно малым.
Исследуемый орган извлекают из организма сразу же после умерщвления животного, чтобы посмертные изменения были минимальными. Срезы делают с помощью лезвия бритвы или микротома, затем переносят в сосуд с подходящей средой суспендирования и изучают их метаболизм и действие введенных соединений на обменные процессы. Тканевые срезы часто исследуют манометрическими методами (разд. 8.6.2); при этом, поскольку срезы бывают недостаточно тонкими, для создания аэробных условий и обеспечения кислородом глубокорасположенных слоев клеток приходится применять газовые смеси, содержащие до 95% кислорода. Недостатком данного метода является то, что внешние слои клеток срезов находятся в среде с токсической концентрацией кислорода.
7. Использование растительного материала
Выбор методов при изучении метаболизма у растений определяется в основном степенью организации растения. Одноклеточные и многоклеточные водоросли, например, хорошо растут на простых, чаще всего неорганических питательных средах при соответствующих внешних условиях. Такие водоросли можно рассматривать как интактные организмы; они имеют относительно простое строение и являются удобным экспериментальным материалом для изучения фундаментальных биохимических процессов, которые трудно исследовать на высокоорганизованных растениях. В качестве классического примера можно привести такие растительные организмы, как Scenedesmus и Chlorella, которые используются для изучения фиксации углекислого газа. Эти системы благодаря удобству контроля за их ростом и простоте поставки экзогенных соединений клеткам особенно удобны для изучения действия на обмен веществ таких факторов, как освещение, температура, питание и т. д.
На более высоких уровнях организации — у высших растений— доставка экзогенных соединений в соответствующий участок внутри растения в значительной степени затруднена. Если растение растет в почве, исследуемое соединение в виде раствора вносят в эту почву, откуда оно затем всасывается корнями. Другой способ состоит в том, что растение извлекают из почвы и корни помещают в раствор исследуемого соединения на определенный период времени. Раствором соединения можно опрыскивать растение или наносить его непосредственно на листья. Для изучения распределения соединения и его метаболитов внутри растительного организма исследуют отдельные его части — корни, побеги, листья, почки и цветы.
Основная трудность, возникающая при изучении метаболизма у растений, заключается в том, что в отличие от тканей животных растительные ткани не содержат достаточно крупных и сложных структур. Отдельные части растения можно изолировать, помещать в соответствующую среду, а затем изучать их метаболизм in vitro. Приготовление срезов, дисков, гомогенатов и выделение клеточных органелл из растительных тканей осуществляют такими же способами, как и из тканей животных.
8. Культуры тканей и клеток
Как мы уже говорили, изучение метаболизма на уровне организма или органа связано с целым рядом трудностей. Иногда это обусловлено еще и тем, что некоторые растения содержат очень мало живой ткани, например ткани меристемы, возникают также трудности доставки соединения в определенный участок растения и контроля за ним. Поэтому выращивание тканей и клеток in vitro имеет ряд преимуществ. В соответствующих экспериментальных условиях можно изучать рост, деление и дифференцию клеток, при этом в значительной степени облегчается доставка соединений к клеткам и тканям и изучение их действия. In vitro можно выращивать большие количества той или иной ткани по сравнению с естественным содержанием ее в интактном организме.
Широкое использование данного метода в эволюционных, физиологических, общемедицинских и фармакологических исследованиях обусловлено тем, что культивирование клеток и тканей позволяет преодолевать многие физические, физиологические и биохимические ограничения, накладываемые сложным строением организма. Метод позволяет изучать потенциал развития клетки, т.е. способность клетки в пределах, обусловленных генотипом, образовывать при соответствующих химических и физических условиях любой другой тип клетки. Несмотря на то что культуры клеток растительных и животных тканей мало чем отличаются друг от друга, клетки растительных тканей могут размножаться в менее сложных средах, чем клетки тканей животных.
Культуры отдельных частей растения, например корня или меристемы, получают, помещая вырезанные части растения в стерильных условиях в питательную среду, поддерживающую их рост и развитие. В таких средах, которые бывают жидкими или полужидкими, можно выращивать также изолированные растительные и животные клетки. Питательные среды имеют довольно сложный химический состав; как правило, они содержат источник углерода (например, сахар), смесь неорганических солей, микроэлементы, витамины и факторы роста.
Часто в культуральные среды вносят сложные питательные добавки: к культурам животных клеток добавляют сыворотку крови, а к культурам растительных — кокосовое молоко. Чтобы обеспечить полную воспроизводимость данных, лучше всего там, где это возможно, пользоваться питательной средой постоянного состава. В подходящей среде в процессе роста и деления клеток образуются группы клеток, на которых и проводят исследование. Если культуру необходимо аэрировать (например, когда она представляет собой жидкую суспензию), коническую колбу или специальный сосуд с суспензией встряхивают или вращают. В тех случаях, когда требуется получить дезагрегированную суспензию, в раствор добавляют определенные ферменты, например трипсин к суспензии животных клеток или пектиназы к суспензии растительных.
Оценку клеточного роста на основе подсчета количества клеток и увеличения клеточного объема, позволяющих судить о размере клеток, удобнее проводить не в интактной ткани, а в клеточной суспензии. Соединение, внесенное в культуральную среду, поступает непосредственно в клетки, что позволяет легко проследить за действием этого соединения на рост и обмен веществ в клетке. Применение этого метода для культивирования растительных клеток с удаленной внешней стенкой (культуры протопластов) и для поддержания роста вирусов еще более расширяет границы его использования.
Клеточные культуры широко используют в микробиологии для получения в большом количестве водорослей, грибов и бактерий для исследовательских целей. Для выращивания микроорганизмов применяют как жидкие, так и твердые среды; по своему химическому составу они, как правило, менее сложны, чем те, которые применяются для выращивания культур клеток животных и высших растений. Скорость роста микроорганизмов во много раз выше, чем скорость размножения других клеточных культур, поэтому поддерживать стерильность культур растительных и животных клеток чрезвычайно трудно — любое заражение культуры микробами приводит к быстрому инфицированию выращиваемой культуры.
Подобно другим методам in vitro, применение тканевых и клеточных культур ставит перед исследователями проблему экстраполяции полученных результатов к целому организму, особенно в тех случаях, когда при культивировании растительные и животные клетки дифференцируются.
9. Фракционирование клеток
Фракционирование клеток состоит из двух последовательных стадий — гомогенизации и разделения. На стадии гомогенизации структура ткани разрушается и ткань превращается в так называемый гомогенат. На второй стадии — разделении — происходит группирование отдельных компонентов гомогената по принципу общности их физических свойств, таких, как размер и плотность.
При идеальных условиях выделения внутриклеточные компоненты можно было бы получать в том же виде и количестве, в которых они существуют в интактных клетках, не нарушая таким образом их морфологической структуры и не изменяя их активности. Однако большинство существующих в настоящее время методов фракционирования всем этим требованиям не удовлетворяет, и при выборе того или иного метода часто приходится иметь в виду, что в ходе фракционирования за счет сохранения морфологической структуры клетки может нарушиться ее активность, и наоборот.
Выбор ткани для фракционирования определяется конкретными условиями эксперимента и объектом исследования. Ткани и клетки различных органов различаются по составу, хрупкости и плотности,- что в свою очередь определяет выбор того или иного метода выделения. Печень, например, является идеальным объектом для изучения функционирования митохондрий, поскольку именно в клетках печени митохондрии содержатся в особенно больших количествах. Ткань тимуса (зобной железы) чаще других тканей используется для выделения ядер, так как ядра тимоцитов составляют до 50% клеточной массы.
Клетки различных тканей обычно гетерогенны по форме и размерам; подобная гетерогенность точно в такой же степени характерна и для выделяемых из гомогенатов тканей субклеточных фракций. В этой связи химический анализ выделенных фракций может дать лишь усредненные данные о составе этих фракций. Различные органы животных отличаются друг от друга и по содержанию в них крови и соединительной ткани: чем больше соединительной ткани содержится в органе, тем хуже ткань поддается гомогенизации и тем труднее поэтому выделить из нее субклеточные компоненты.
9.1 Приготовление гомогенатов тканей и клеток
Гомогенизация приводит к потере морфологических и биохимических свойств, характерных для данной ткани. Такая потеря не существенна, если гомогенизация проводится как предварительная стадия выделения из ткани какого-либо химического соединения.
Однако в тех случаях, когда изучают метаболические процессы, морфологическая и биохимическая целостность ткани должна быть сохранена в максимальной степени. Целью гомогенизации, которая, к сожалению, по-прежнему остается эмпирическим методом, является разрушение тканей, клеточных стенок и (или) мембран и высвобождение клеточного содержимого. Для этого применяются самые разнообразные методы и приборы, хотя лежащие в их основе принципы не всегда ясны. Лишенная прочной теоретической базы и необходимого арсенала стандартных методов, гомогенизация представляет собой скорее искусство, чем науку. В силу того, что различные ткани в значительной степени отличаются одна от другой как по хрупкости определенных клеточных органелл, так и по устойчивости клеток и тканей к разрушению, при гомогенизации любого биологического материала всякий раз неизбежно возникают специфические проблемы, которые можно разрешить только путем проб и ошибок. В основном гомогенизация применяется как стадия, предшествующая разделению клеточных компонентов, которая дает возможность установить внутриклеточную локализацию метаболических процессов. Гомогенаты успешно используются и при изучении поглощения и метаболизма соединений в тех случаях, когда доставка их в интактные клетки затруднена в силу недостаточной проницаемости мембран.
9.2 Выбор среды суспендирования
Объективных критериев для выбора той или иной среды суспендирования при гомогенизации не существует. Некоторые рекомендации можно почерпнуть из литературы, однако окончательный выбор всегда зависит от результатов предварительных опытов с применением различных сред.
Обычно для создания в среде необходимого осмотического давления, предохраняющего частицы от набухания и разрыва, применяют сахарозу. Если сахароза затрудняет исследование свойств ферментов, ее заменяют маннитом. Существует целый ряд индивидуальных прописей по сохранению целостности частиц и защите ферментов от инактивации. Рекомендуемые растворы различаются по концентрации сахарозы или присутствию таких веществ, как ЭДТА, глутатион, р-меркаптоэтанол и т. д. Иногда вместо солевых растворов используют неионные среды, так как, например, в гомогенатах печени солевые растворы вызывают агглютинацию полиморфно-ядерных лейкоцитов и органелл. При работе с гомогенатами селезенки, Наоборот, сахароза (0,25 М) обладает более выраженным агглютинирующим действием, чем КС1 (0,2 М).
Для выделения ядер и хромосом пользуются лимонной кислотой, которая обладает способностью подавлять активность «нейтральных» дезоксирибонуклеаз.
Для выделения ядер применяют растворы глицерина и этиленгликоля, а для выделения пластид из клеток растений — карбоваксы (полимеры этиленгликоля). Хлоропласты обычно выделяют в средах, содержащих не сахарозу, а маннит и сорбит.
Анализ ферментов в растительных экстрактах иногда значительно усложняется в силу того, что в процессе гомогенизации выделяется большое количество фенолов, которые образуют водородные связи с карбонильными группами, участвующими в образовании пептидных связей белков, а это, по-видимому, вызывает инактивацию многих ферментов. Во избежание этого к экстрактам добавляют поливинилпирролидон, образующий с фенолами нерастворимый комплекс, который затем удаляют из экстракта фильтрованием.
Для выделения субклеточных органелл можно применять неводные среды. Суспендирующая среда в этом случае представляет собой смесь легкого и тяжелого органических растворителей, например смесь эфира с хлороформом или бензола с четыреххлористым углеродом. Плотность среды можно изменять таким образом, чтобы при последующем центрифугировании исследуемые частицы либо всплывали на поверхность, либо осаждались. «Неводное» фракционирование применяют для выделения хлоропластов и лейкоцитов, а также гранул гемосидерина из селезенки. Недостатками этого метода являются нарушение морфологической структуры некоторых видов ткани и инактивация отдельных ферментов органическими растворигелями.
9.3 Способы разрушения тканей и клеток
Для разрушения клеток чаще всего применяют физические методы. Большинство животных клеток разрушается сравнительно легко, однако при разрушении растительных и бактериальных клеток зачастую приходится сталкиваться со значительными трудностями, связанными с наличием клеточных стенок. Физические методы разрушения клеток подразделяются в зависимости от того, происходит ли оно под действием сил трения между клетками и твердыми веществами (растирание клеток с твердыми материалами) или гидродинамически (разрушение клеток в жидких средах).
Растирание клеток с твердыми материалами. В современной модификации этот метод состоит в растирании клеток с песком или абразивным порошком в ступке при помощи пестика. В настоящее время благодаря появлению более мягких способов разрушения этот метод применяется для разрушения животных клеток довольно редко, однако им по-прежнему пользуются для разрушения растительных и бактериальных клеток. Желательно, чтобы абразивные частицы были как можно более острыми и имели такой же размер, что и разрушаемые клетки. Недостаток метода заключается в том, что при разрушении клеток может нарушаться структура наиболее крупных органелл, таких, например, как хлоропласты.
Хорошие результаты дает продавливание клеток, смешанных с абразивными частицами, через пресс Хьюза. Влажные клетки с абразивными частицами помещают в трубку при температуре около —5°С, а затем однократным ударом по поршню, создающим скачкообразное изменение давления, проталкивают клеточную массу через узкое отверстие диаметром около 0,25 мм. Модификацией этого метода является продавливание клеток при температуре —25°С; роль абразивных частиц выполняют в этом случае кристаллы льда. Чтобы добиться максимального разрушения бактериальных клеток, приходится иногда повышать давление до 5,5-107 Па.
Клетки бактерий можно разрушать также и путем механического встряхивания суспензий частиц с абразивным порошком с частотой 300—3000 колебаний в минуту при помощи встряхивателя Микля, в который добавляются мелкие стеклянные бусинки диаметром от 50 до 500 мкм. Однако возникающая при встряхивании сильная вибрация часто вызывает разрушение клеточных органелл.
Разрушение клеток в жидких средах. Разрушение клеток, находящихся в суспензии, происходит либо при вращении лопастей или поршня (блендеры), либо при поступательном движении вверх и вниз поршня или шаров (гомогенизаторы).
Блендеры, как правило, имеют режущие лопасти, вращающиеся с большой скоростью. Количество и конструкция этих лопастей бывают разными, но все они обычно заострены под прямым углом друг к другу, а форма их обеспечивает хорошее перемешивание содержимого сосуда. Суспензию клеток помещают в специальный стакан, который имеет по всей высоте раструбы и в поперечном сечении выглядит как клеверный лист. Для поддержания низкой температуры в процессе гомогенизации стакан помещают в лед. Благодаря особому расположению лопастей и конструкции стакана в ходе фракционирования возникают гидродинамические силы. Метод достаточно универсален и широко применяется для фракционирования клеток, однако следует иметь в виду, что при быстром вращении лопастей гомогенизаторов возникают некоторые нежелательные эффекты.
Большинство гомогенизаторов преставляют собой прибор, состоящий из пестика с ручным (гомогенизаторы Даунса и Тёнбрэка) или механическим (гомогенизатор Поттера—Эльвегёйма) приводом, который вращается или движется вверх и вниз в стеклянном цилиндрическом сосуде. Необходимо следить за тем, чтобы зазор между пестиком и стенками сосуда оставался постоянным, так как скорость разрушения клеток зависит не только от скорости вращения пестика, но и от соотношения между радиусами пестика и сосуда. Сосуд закрепляется неподвижно,- поэтому скорость вращения суспензии изменяется от минимальной у его стенок до максимальной у поверхности пестика; следовательно, чем меньше расстояние между этими поверхностями, тем выше градиент скорости. Возникающие при высоких скоростях силы достаточны для разрушения довольно тонких мембран животных клеток; растительные и бактериальные клетки при этом не разрушаются.
Эффективность гомогенизации в значительной степени зависит от наличия в измельчаемом материале сосудистой и соединительной ткани, для удаления которой ткань перед гомогенизацией пропускают через специальную мясорубку с отверстиями диаметром 0,88 мм8 .
Полиморфоядерные лейкоциты разрушаются более мягкими методами— при помощи пипетки. Эозинофилы разрушают путем быстрого пропускания под давлением через мелкорешетчатое сито.
Разрушение клеток с помощью высокого давления. Этот метод применяется в основном для разрушения микробных клеток. Для этой цели пользуются специальными прессами, например френч-прессом (French Pressure), в котором создается давление до 10,4-107 Па.
Суспензию клеток загружают в камеру из нержавеющей стали (рис. 1.4) при закрытом положении игольчатого клапана, посредством которого камера сообщается с внешней средой. Затем камеру переворачивают, открывают клапан и поршнем вытесняют из камеры воздух, после чего клапан снова закрывают, а камеру возвращают в исходное положение и устанавливают на неподвижном основании. С помощью гидравлического пресса создается требуемое давление на поршень; по достижении в камере определенного давления игольчатый клапан немного открывается, давление в камере несколько уменьшается, и в этот момент клетки лопаются. Вытекающую из выходного отверстия камеры клеточную массу собирают при открытом положении игольчатого клапана, поддерживая в камере постоянное давление. К сожалению, неизвестно, какие именно силы возникают в камере и как им противостоят клетки и клеточные компоненты. Разрушение с помощью ультразвука. Клетки можно разрушить также с помощью высокочастотных ультразвуковых колебаний. Механизм такого разрушения окончательно не выяснен, однако установлено, что при обработке клеточных суспензий ультразвуком в среде создается высокочастотное изменение давления. Основным недостатком данного метода является то, что в процессе обработки ультразвуком выделяется значительное количество тепла. Чтобы избежать разогревания, сосуд с суспензией помещают в лед; конструкция сосуда такова, что жидкость непрерывно циркулирует и охлаждается у стенок. Некоторые сосуды помещают в холодильные камеры; однако далеко не всегда удается устранить местное нагревание.
Прочие методы. К ним относятся: разрушение клеток методом осмотического шока, переваривание клеточных стенок ферментами, например лизоцимом, и сложными ферментными препаратами, выделенными из улиток и содержащими целлюлазу, хитиназу и липазу. Для разрушения клеток некоторых видов успешно применяют замораживание и оттаивание, автолиз и обработку органическими растворителями, такими, как этилацетат и толуол.
Как мы уже говорили, в большинстве случаев разрушение клеток сопровождается выделением тепла. Ввиду того что при высоких температурах многие ферменты инактивируются, все процедуры по разрушению клеток и выделению клеточных органелл следует проводить при пониженных температурах. Для этого все работы проводят в холодных комнатах при температуре около 0°С или охлаждают клеточные суспензии с помощью льда. Следует отметить, что некоторые ферменты неустойчивы к холоду и при охлаждении также теряют свою активность.
При гомогенизации разного рода биологических тканей возникает множество частных проблем, которые разрешаются в основном путем проб и ошибок. Достаточную воспроизводимость результатов можно получить лишь при тщательном контроле за такими параметрами, как температура, продолжительность и скорость разрушения клеток, а также применяемое рабочее давление. Идеальным считается такой гомогенат, который легко поддается дальнейшему фракционированию.