| Похожие рефераты | Скачать .docx | Скачать .pdf |
Курсовая работа: Получение рекомбинантного аденовируса CELO
Получение рекомбинантного аденовируса CELO
СОДЕРЖАНИЕ
Введение
1. Литературный обзор
1.1 Генетические механизмы наследственности
1.1.1 Характеристика нуклеиновых кислот
1.1.2 Нуклеозиды и нуклеотиды
1.1.3 Первичная структура ДНК
1.1.4 Вторичная структура ДНК
1.1.5 Третичная структура (суперспирализация) ДНК
1.2 Эпигенетические механизмы наследственности
1.2.1 Контроль активации генов
1.2.2 Роль хроматина в жизни клетки
1.2.3 Структура хроматина
1.2.4 Характеристика «гистонового кода»
1.3 Современные методы ДНК-технологии
1.3.1 Содержание генетической инженерии
1.3.2 Методы исследования структуры и функции гена
1.3.3 Предмет рекомбинантной ДНК-биотехнологии
1.3.4 Получение чужеродной ДНК
1.3.5 Конструирование рекомбинантных ДНК
1.3.5.1. ДНК-рестриктазы и ДНК-метилазы
1.3.5.2 ДНК-лигазы
1.3.6 Векторы
1.3.7 Введение вектора в клетку
1.3.8 Обнаружение рекомбинантного клона
2. Научно-исследовательская часть
2.1 Характеристика объекта исследования
2.2 Описание эксперимента
2.3 Обработка экспериментальных данных
Заключение
Проблема изучения структуры и функции ДНК, а также возможности ее искусственного изменения и создания организмов с заданными наследственными свойствами - одна из центральных задач биотехнологии.
Предлагаемая курсовая работа структурно состоит из двух частей: первая содержит литературный обзор наиболее важных положений учения о ДНК и характеризует современное видение проблемы. Вторая часть научно- исследовательская, выполненная автором на базе Государственного НИИ эпидемиологии и микробиологии им. Н.Ф. Гамалеи (г. Москва) с целью получения плазмиды, пригодной для создания рекомбинантных аденовирусов.
Первая часть выполнена под руководством профессора Н.С. Колобелина (Воронежская государственная биофабрика), вторая часть под руководством канд. биологических наук, старшего научного сотрудника Д.Ю. Логинова (ГУНИИ ЭМ им. Гамалеи, г. Москва).
25 апреля 1953 г. в журнале «Nature» были напечатаны две заметки. Авторы первой из них, названной «Структура дезоксирибонуклеиновой кислоты», - 25-тилетний американский биохимик Джеймс Уотсон и 37-тилетний английский биофизик Фрэнсис Крик на полутора страницах описали знаменитую двойную спираль и попытались объяснить, как молекуле ДНК удаётся добиться такой точности самовоспроизведения.
В другой статье ровесник, соотечественник и коллега Крика Морис Уилкинс с соавторами опубликовал высококачественные рентгенограммы молекулы ДНК, которые подтверждали гипотезу её «спиральности».
Ради справедливости следует отметить, что к открытию структуры ДНК имела прямое отношение Розалинда Франклин, которая в лаборатории М. Уилкинса сделала прекрасные рентгенограммы РНК, которые были переданы Д. Уотсону и Ф. Крику.
Указанное открытие инициировало целый ряд исследований в области наук о жизни. Начиная с момента открытия, а особенно после расшифровки генома человека научные изыскания дают значительные практические результаты, наиболее перспективные из которых следующие: создание организмов-продуцентов биологически активных веществ; разработка генетических вакцин; развитие генной терапии; применение трансгенных растений и животных для интенсификации сельского хозяйства.
1.1 Генетические механизмы наследственности
1.1.1 Характеристика нуклеиновых кислот
Нуклеиновые кислоты - это природные высокомолекулярные органические соединения, обеспечивающие хранение и передачу наследственной информации в живых организмах.
Нуклеиновые кислоты - высокомолекулярные соединения со строго определённой линейной последовательностью мономеров. Структура ДНК и РНК - способ «записи информации», обеспечивающий формирование в организме двух информационных потоков. Один из потоков осуществляет воспроизведение информации, заключённой в молекулах ДНК. Удвоение молекул ДНК называется «репликацией». В результате этого процесса и последующего деления дочерние клетки наследуют геном родительской клетки, в котором содержится полный набор генов, или инструкции о строении РНК и всех белков организма. Второй поток информации реализуется в процессе жизнедеятельности клетки. В этом случае происходит «считывание», или транскрипция генов в форме полинуклеотидных последовательностей гпРНК и использование их в качестве матриц для синтеза соответствующих белков. В последнем случае осуществляется «перевод» (трансляция) информации, заключённой в тРНК, на «язык» аминокислот. Этот поток информации от ДНК через РНК на белок получил название «центральная догма биологии». Суть новой молекулярной биологии заключается в следующем: всё начинается с ДНК и заканчивается белком. Между ДНК и белком находится РНК, которая является переносчиком генетической информации от ДНК к белкам. Дальше генетическая информация переносится от РНК в особые частицы рибосомы, где реализуется при биосинтезе белков. Механизм характерен для всех живых организмов, за исключением некоторых РНК-содержащих вирусов.
ДНК и РНК состоят из мономерных единиц - нуклеотидов, поэтому нуклеиновые кислоты называют полинуклеотидами.
Строение нуклеотидов. Каждый нуклеотид содержит 3 химически различных компонента: гетероциклическое азотистое основание, моносахарид (пентозу) и остаток фосфорной кислоты. В состав нуклеиновых кислот входят азотистые основания двух типов: пуриновые - аденин (А), гуанин (G) и пиримидиновые - цитозин (С), тимин (Т) и урацил (U) ~ см. рисунок 1.1.
Пентозы в нуклеотидах представлены либо рибозой (в составе РНК), либо дезоксирибозой (в составе ДНК). Чтобы отличить номера атомов в пентозах от нумерации атомов в основаниях, к цифре добавляют штрих (') – 1’, 2', 3', 4' и 5'. Пентозу соединяет с основанием N-гликозидная связь, образованная Сг атомом пентозы (рибозы или дезоксирибозы) и N1-атомом пиримидина или N9- атомом пурина.

Нуклеозиды - соединения, состоящие из остатков азотистого основания и углевода - рибозы или дезоксирибозы.
Нуклеотид - это нуклеозид, к которому присоединены молекулы фосфата - см. рисунок 1.2.

Остов нуклеиновой кислоты имеет одинаковое строение по всей длине молекулы и состоит из чередующихся групп - пентоза - фосфат - пентоза. Уникальность структуры и функциональная индивидуальность молекул ДНК и РНК определяется их первичной структурой - последовательностью азотистых оснований в полинуклеотидной цепи /4/.
1.1.3 Первичная структура ДНК
Первичная структура ДНК - порядок чередования дезоксирибонуклеозидмонофосфатов в полинуклеотидной цепи. Каждая фосфатная группа в полинуклеотидной цепи, за исключением фосфорного остатка на 5 '-конце молекулы, участвует в образовании двух эфирных связей с участием 3' и 5 '-углеводных атомов двух соседних дезоксирибоз, поэтому связь между мономерами обозначают 3',5' -фосфодиэфирной.
Концевые нуклеотиды ДНК различают по структуре: на 5'-конце находится фосфатная группа, а на 3'-конце цепи - свободная ОН-группа. Эти концы называют 5' и 3'-концами — см рисунок 1.3.
Линейная последовательность дезоксирибонуклеотидов в полимерной цепи ДНК обычно сокращенно записывают с помощью однобуквенного кода, например A-G-C - Т- Т- А- С- А от 5'-к З '-концу.

В каждом мономере нуклеиновой кислоты присутствует остаток фосфорной кислоты. При рН 7 фосфатная группа полностью ионизирована, поэтому in vivo нуклеиновые кислоты существуют в виде полианионов (имеют множественные отрицательный заряд). Остатки пентоз тоже проявляют гидрофильные свойства. Азотистые основания почти нерастворимы в воде, но некоторые атомы пуринового и пиримидинового циклов способны образовывать водородные связи.
Молекула ДНК имеет форму спирали, образованную двумя полинуклеотидными цепями, закрученными относительно друг друга и вокруг общей оси. Двойная спираль правозакрученная, полинуклеотидные цепи в ней антипараллельны, т.е. если одна из них ориентирована в направлении 3' —> 5', то вторая в направлении 5'—> 3'. Поэтому на каждом из концов молекулы ДНК расположены 5' -конец одной цепи и 3'-конец другой цепи.
Двойная спираль стала символом новой, молекулярной биологии. Спираль построена за счёт комплементарных оснований. Именно двухцепочечностью и комплементарностью оснований объясняются биологические функции ДНК. В правовинтовой спирали две водородные связи в паре А - Т и три водородные связи в паре G - С . Естественно, последняя пара прочнее, а первая слабее. Модель Уотсона Крика объяснила правило Чаргаффа - большое основание - пуриновое дополняется маленьким основанием - одиночным гетероциклом пиримидина. То же самое вG-CивA-T. Благодаря этому размеры пар одинаковы по ширине, что и приводит к регулярной двойной спирали. Спираль не деформируется, поскольку то, что её скрепляет внутри, симметрично по отношению друг к другу, хотя и повёрнуто. Плоскости азотистых оснований наклонены по отношению к оси спирали образуя третичную структуру. В местах сгиба двойная спираль имеет изломы, то есть нарушается классическая двойная структура ДНК.
1.2 Эпигенетические механизмы наследственности
1.2.1 Контроль активации генов
Существует мнение, что все свойства клетки и даже многоклеточного организма однозначно определены последовательностью нуклеотидов в ДНК. В настоящее время это положение подвергается критике. Выдвинуты представления об эпигенетических механизмах избирательной экспрессии генов, ведущая роль в которых принадлежит хроматину. Именно на уровне хроматина работают основные механизмы программирования развития живого организма. Эти механизмы базируются на структурно-функциональных особенностях нуклеосомы, гистонов, формирующих нуклеосомную сердцевину, межнуклеосомных взаимодействиях, тонко регулируемых внешними сигналами. На уровне нуклеосомы реализуется «гистоновый код», управляющий сложнейшим каскадом биохимических реакций в клетке. Все вышеуказанные механизмы обеспечивают важнейшее приспособление организмов к изменяющимся условиям внешней и внутренней среды.
Геном каждой клетки человека составляет около 40 тысяч генов, но все они одновременно не используются. В каждом клеточном типе работает около 20 тысяч генов. Как минимум половина всех работающих генов необходима для поддержания жизнедеятельности любой клетки, другая половина определяет специализацию клетки, т.е. будет ли она принадлежать печени, почкам, селезёнке, другому органу или ткани. Все клетки содержат одинаковую информацию, записанную в молекуле ДНК, но при развитии организма эта информация в тканях или органах считывается избирательно, что и приводит к огромному разнообразию клеток в организме.
Существует несколько уровней контроля активации генов. Один из них - модификация ДНК, соответствующая тем генам, которые необходимо выключить, т.е. этот ген должен молчать в данном типе клеток. При этом данный признак сохраняется и при удвоении клетки. Однако ДНК сама по себе несёт важнейшую функцию кодирования аминокислотной последовательности белков, и человек не может произвольно её модифицировать, не затронув этой информации и не изменив способность ДНК к удвоению.
На сегодня единственной изученной модификацией ДНК, которая не нарушает кодирование и копирование, является метилирование цитозина, которое выключает ген и он становится молчащим, и, главное, это свойство закрепляется в наследственности, т.е. в дочерней клетке сохраняется информация о том, что данный ген должен молчать.
Как же эта модификация влияет на активацию гена - на получение РНК- копии данного гена, служащей в дальнейшем матрицей для синтеза белка?
Указанная модификация копируется при удвоении ДНК. Возможны два механизма контроля активации гена. Первый - отталкивание белков, активирующих ген, второй - привлечение к метилированной ДНК белков, которые селективно узнают только метилированную ДНК и участвуют в очень плотной её упаковке за счёт изменения структуры хроматина - формы существования генома высших организмов. Именно на уровне хроматина происходит основной контроль активации генов.
Благодаря расшифровке генома возникли две новые науки: геномика, изучающая геномы, и протеомика, изучающая совокупность клеточных белков. Почти 50 лет после открытия ДНК господствовал постулат: один ген синтезирует один белок. После расшифровки генома появилась новая догма: один ген - много белков. Установлено, что гены у высших организмов состоят из кодирующих и некодирующих участков. В результате комбинаторики из одного гена путём соединения кодирующих участков можно получить очень много белков. Если у человека около 40 тысячи генов (по геномике), то по протеомике можно получить минимально более 100 тысяч белков. Это только на уровне первичной структуры, фактическая ситуация ещё сложнее, потому что белки подвергаются модификациям, на них «навешивается» огромное количество разных групп, что приводит к колоссальному разнообразию. Если геном постоянен, то протеом очень вариабелен. Уотсон очень много сделал для расшифровки генома человека, которая закончилась в 2003 году /3/.
1.2.2 Роль хроматина в жизни клетки
Хроматин - нуклеопротеид, комплекс белков с ДНК и РНК в ядрах клеток, обеспечивает упаковку сравнительно длинного фрагмента ДНК в значительно меньшее по размерам ядро. Хроматин - чрезвычайно сложный и динамичный комплекс и в живых клетках высших организмов он служит носителем генетической информации.
Избирательное и комбинаторное извлечение информации из ДНК, происходящее на уровне хроматина, приводят к огромному разнообразию фенотипического проявления наследственного материала. Неудивительно поэтому, что при почти 80 %-ной схожести геномов человека и мыши - эти два организма весьма существенно отличаются друг от друга.
На уровне хроматина, а не ДНК наиболее эффективно работают многие ферменты, участвующие в избирательном считывании генетической информации, её удвоении при делении клетки и исправлении возникающих при этом ошибок
1.2.3 Структура хроматина
Универсальные компоненты хроматина - белки гистоны.
Гистоны - это небольшие по молекулярной массе белки с высоким содержанием положительно заряженных аминокислотных остатков - лизинов и аргининов, локализованных преимущественно в N- и С-концевых неструктурированных областях белка, так называемых «хвостах», которые играют главную роль в эпигенетических механизмах. В центральных, наиболее консервативных участках гистонов преобладают гидрофобные аминокислотные остатки, необходимые для белок-белкового узнавания. Существуют пять типов гистонов: HI, Н2А, Н2В, НЗ, Н4. Последовательность расположения гистонов в нуклеосоме одинакова для организмов-эукариотов всех царств живой природы: грибов, растений, животных.
Рассмотрим строение нуклеосомы. Нуклеосома состоит из восьми молекул комплекса гистонов: по два гистона Н2А и Н2В с высоким содержанием лизина, по два богатых аргинином гистона НЗ и Н4. Этот гистоновый октамер называется «кор» (от лат. cor - сердцевина) На кор навивается ДНК в виде левозакрученной суперспирали с шагом 28 А, содержащей 80 пар нуклеотидов на виток. Всего на кор в нуклеосоме навивается 1,75 витка или 146 пар нуклеотидов. Описанная нуклеосома носит название «минимальной» или «кор-нуклеосомы». Если в состав нуклеосомы входит гистон HI, который связывается с межнуклеосомной ДНК, или линкером, то такую нуклеосому называют полной; на неё навивается 2-2,5 витка ДНК (160-200 пар нуклеотидов).
В основе организации нуклеосомы лежит уровень многоуровневого узнавания. На первом уровне гистоны попарно узнают друг друга. Молекула каждого гистона состоит из центрального структурированного трёхспирального домена (одного длинного и двух коротких альфа-спиральных участков, соединённых петельным сегментом, так называемая гистонная укладка) и двух неструктурированных N и С-«хвостов». Спиральные домены гистонов взаимодействуют друг с другом («рукопожатие»), в результате чего образуются гетеродимеры НЗ - Н4 и Н2А - Н2В. Эти димеры образуют тетрамер НЗ - Н4 и два димера Н2А - Н2В, формирующие гистоновый октамер, который имеет клинообразную форму, обеспечивающую левое закручивание спирали ДНК вокруг себя. На его поверхности, обращённой к ДНК, формируются белковые структуры {мотивы), которые можно разделить на три основных типа.
Мотивы первого типа образованы спаренными петельными сегментами гистоновых димеров (бета-мосты).
Мотивы второго представляют собой спаренные N-концевые сегменты первых спиральных доменов каждого гистона в гетеродимерах. К третьему типу относят два мотива, которые формируются двумя дополнительными альфа спиральными участками гистона НЗ, расположенными по концам суперспирали
ДНК, где ДНК входит и покидает иуклеосому. Суммарное число всех мотивов равно 14, что соответствует числу витков двойной спирали ДНК, накрученной вокруг октамера.
Все эти мотивы содержат положительно заряженные аминокислоты (в основном аргинины), что обеспечивает электростатический контакт октамера с отрицательно заряженными фосфатами сахарного остова ДНК. При этом 14 малых бороздок ДНК втягивают в себя 14 аргинин содержащих мотивов гистонового октамера, расположенных на поверхности гистонового кора и обращенных к ДНК. Это приводит к жёсткому фиксированию ДНК, которое в малой степени зависит от её нуклеотидной последовательности, что и обеспечивает универсальность компактизации ДНК на нуклеосомах. Консервативная глобулярная часть гистонов участвует в формировании гистонового октамера и фиксировании на нём молекулы ДНК. Так обеспечивается первый уровень компактизации. При этом ДНК оказывается на поверхности кора и остаётся доступной для взаимодействия с другими белками. В последние годы изучалась роль гистоновых хвостов.
Согласно рентгеноструктурному анализу нуклеосом, суперспираль ДНК в нуклеосоме закручена так, что обеспечивается выход «хвостов» гистонов на поверхность нуклеосомы. «Хвост» гистона НЗ простирается далеко от места выхода на
поверхность нуклеосомы и фиксируется межнуклеосомными контактами. «Хвост» гистона Н4 имеет много контактов с поверхностью димера Н2А - Н2В соседней нуклеосомы. «Хвосты» гистонов выходят на поверхность хроматиновой фибриллы, участвуют в межнуклеосомном взаимодействии, очень подвижны и подвергаются многочисленным модификациям: ацетилированию, фосфорилированию, метилированию, убиквитинилированию и АДФ-рибозилированию. Эти модификации приводят к изменению заряда, гидрофобности и других свойств поверхности белковых глобул. В результате формируется сложная матрица для узнавания её другими регуляторными белками и внешними сигналами. Поскольку концевые домены гистонов участвуют и в межнуклеосомном взаимодействии, вышеперечисленные модификации влияют и на характер упаковки хроматиновой фибриллы, разрыхляя или, наоборот, уплотняя её, что, в свою очередь облегчает или затрудняет доступ к ДНК многочисленным регуляторным факторам.
Эти свойства «хвостов» гистонов в структуре нуклеосомы имеют большое значение для расшифровки и понимания механизмов функционирования хроматина, его поведения при активации генов, репрессии их и многих других процессов, связанных с доступом к ДНК /4/.
1.2.4 Характеристика «гистонового кода»
Наиболее разработанной в настоящее время моделью функционирования хроматина считается «гистоновый код». Это - разнообразный набор модификаций гистоиовых «хвостов» на поверхности нуклеосом, который можно целенаправленно менять и передавать по наследству. Он и определяет функциональное состояние гена.
«Гистоновый код», по-видимому, является основным эпигенетическим механизмом, контролирующим включение или выключение генов и передачу программы этого контроля по наследству от клетки к клетке.
«Гистоновый код» оказался идеальным эпигенетическим механизмом, с помощью которого можно писать программу каскадного включения - выключения генов при развитии, не затрагивая информацию о белках, записанную на самой ДНК.
Гипотеза «гистонового кода» предполагает, что маркировки на модифицированных концевых заменах гистонов должны узнаваться регуляторными белками. Первым был обнаружен так называемый бромодомен, который специфически узнает ацетилированные лизины на гистоновых «хвостах». Бромодомен присутствует во многих регуляторах транскрипции генов (например, белок TAFn250). (Субъединица базального транскипционного фактора TFIID) узнаёт, например, комбинацию нескольких ацетильных групп и имеет две последовательно расположенные копии бромодомена, каждая их которых узнаёт свою ацетильную группу.
Другой белковый домен, узнающий маркировки на гистоновых «хвостах», называется хромодоменом. Из несущих хромодомен белков лучше всего изучен НР1 (от англ. Heterochromatin protein). Этот белок принимает участие в структурной организации гетерохроматина - области хроматина, находящейся в высококонденсированном состоянии, где все гены репрессированы. Таким образом, белок НР1 - это маркёр неактивного состояния хроматина. Он имеет высокое сродство к метилированному 9-му лизину (Tys-9) гистона НЗ. Связываясь с метилированным Tys-9 НЗ в одной нуклеосоме, белок HP 1 в свою очередь, метилирует соседние нуклеосомы, что вызывает распознание области метилирования хроматина. Этим достигается эффект высококонденсированного состояния на протяжённых участках хроматина. В настоящее время ведётся поиск доменов, узнающих другие модификации «хвостов» гистонов (фосфорилирование, убиквитинилирование и АДФ-рибозилирование).
Существуют несколько вариантов реализации «гистонового кода».
Согласно одному из них, модификации гистоновых «хвостов», участвующих в межнуклеосомных взаимодействиях, изменяют их суммарный заряд, за счёт чего уменьшается степень компактизации хроматиновой фибриллы, и ДНК становится доступной для регуляторных белков. В этом случае роль ферментов, модифицирующих «хвосты» гистонов, заключается только в увеличении зоны модификаций, и как следствие - усиление эффекта компактизации либо декомпактизации.
Другой механизм изменения статуса гена, маркированного определёнными сигналами по «хвостам» гистонов, связан с факторами перестройки хроматина. В 1992 году на дрожжах впервые было показано, что в присутствии АТФ определенные белковые комплексы обладали способностью изменять структуру хроматина, удаляя или сдвигая нуклеосомы с регуляторных участков ДНК. При этом активировались молчащие гены. В состав такого комплекса входят белки, несущие бромодомены, хромодомены, избирательно узнающие модифицированные «хвосты» гистонов. Это позволяет целенаправленно доставлять комплекс к регуляторному участку гена, где и происходят структурные изменения в хроматине, облегчающие доступ к ДНК белкам, активирующим транскрипцию этого гена.
Получены данные, свидетельствующие, что белки, узнающие «гистоновый код», выполняют транспортную функцию: доставляют к необходимым генам комплекс регуляторных белков, порождающих целый каскад биохимических реакций в области хроматина, который соответствует регуляторному участку гена. В состав комплекса могут входить как модифицирующие белки, так и транскрипционные факторы.
Изучение эпигенетических механизмов уже перестало быть только проблемой фундаментальной науки. Известно, что уровень и характер метилирования ДНК отклоняется от нормы при канцерогенезе и старении. Понимание того, что метилирование ДНК определяется эпигенетическим молчанием хроматина, проливает свет на механизмы этих процессов. Расшифровка специфического эпигенетического «ракового кода» и «кода старения» на уровне метилирования ДНК имеет огромное практическое значение. Итак, сделаем вывод: многочисленные модификации гистоновых «хвостов», экспонированные на поверхности хроматиновой фибриллы, и их комбинации, огромный набор специализированных ферментов, узнающих эти комбинации, позволяют записать очень сложную программу последовательности включения - выключение генов. В этом случае генетической информации, записанной на ДНК, оказывается недостаточно для получения полноценного организма. К информации должна прилагаться инструкция по её использованию. Подтверждением вышесказанного могут служить огромные сложности при получении клонов животных. Если берётся ДНК из клетки любого органа и переносится в яйцеклетку, где всё подготовлено к тому, чтобы дать развитие новому организму, то выясняется, что эпигенетические механизмы развития в исходной клетке давно запущены, стереть эту информацию невозможно и многие гены уже никогда не смогут начать работать. Несмотря на то, что имеется полная генетическая копия, то есть геном представлен абсолютно такой же ДНК, как запустить программу развития, клетка не знает. Этой информацией обладают только ДНК половых клеток и так называемых эмбриональных стволовых клеток - первые деления оплодотворённой яйцеклетки. Только на этой стадии эпигенетические механизмы ещё не включены, и клетка может стать и клеткой печени, и клеткой мозга, и клеткой кожи. На начальных этапах исследования ДНК казалось, что, расшифровав полную генетическую информацию геномной ДНК, появится возможность управлять внутриклеточными процессами. Однако выясняется, что важна и эпигенетическая информация, язык которой гораздо сложнее и от полной расшифровки которой наука ещё очень далека. 1.3. Современные методы ДНК-технологии.
1.3.1 Содержание генетической инженерии
В предыдущих разделах описывались генетические и эпигенетические механизмы наследственности. Современные методики позволяют модифицировать наследственную информацию и конструировать организмы с заданными наследственными свойствами. Эти методы составляют предмет генетической инженерии.
Генетическая инженерия - это искусство использования знаний, методов и техники физико-химической биологии и молекулярной генетики для конструирования организмов с заданными наследственными свойствами. Конечная цель состоит в получение рекомбинантной ДНК с последующим включением её в реципиентную клетку или в осуществлении переноса целых хромосом от клеток-доноров в клетки-реципиентов. В основу генно- инженерных методов заложена способность ферментов (рестриктаз) расщеплять ДНК на отдельные нуклеотидные последовательности, которые могут быть использованы для встраивания их в геномы бактериальных плазмид и фагов с целью получения гибридных форм, состоящих из собственной ДНК и дополнительных встроенных фрагментов несвойственной им ДНК. Таким способом добиваются клонирования генов, когда выделяют нужный отрезок ДНК из какого-либо биообъекта и затем получают любое количество его, выращивая колонии генетически идентичных клеток, содержащих заданный участок ДНК. Другими словами, клонирование ДНК—это получение её генетически идентичных копий.
Генетическую инженерию подразделяют на генную инженерию, геномную инженерию, и хромосомную инженерию. Сущность первой состоит в целенаправленном использовании перестроек естественного генома, осуществляемых in vivo или in vitro, для изменения генетических характеристик известных вирусов и клеток. Примером генной инженерии in vitro является создание молекулярных химер из фрагментов ДНК разного происхождения, включения их в реципиентные клетки Е. coli, Вас. subtilis и др., с последующим культивированием этих организмов в целях получения необходимых белковых продуктов (пептидных гормонов, ферментов и т.д.)
Сущность геномной инженерии заключается в целенаправленной глубокой перестройке генома прокариот или эукариот вплоть до создания новых видов. При геномной инженерии добиваются внесения большого количества дополнительной генетической информации и в результате получают гибридный организм, отличающийся от исходного по ряду признаков.
В случае переноса изолированных хромосом от клетки-донора одного организма в клетку-реципиент другого организма, говорят о хромосомной инженерии. Благодаря хромосомной инженерии стали возможными получение высокомолекулярных БАВ, присущих человеку, лечение наследственных заболеваний, селекция пород домашних животных и различных видов растений.
1.3.2 Методы исследования структуры и функции гена
Образование новых сочетаний генов и их частей в природных условиях, по-видимому, не носит целенаправленного характера, и лишь жесткая проверка естественным отбором может оценить жизненную значимость таких преобразований геномов. Однако сознательное использование в лабораторных условиях основных генетических принципов, лежащих в основе природных перемещений генов, позволило разработать более эффективные системы передачи генетической информации между организмами и приступить к беспрецедентным по информативности исследованиям генетических явлений на молекулярном уровне.
Необходимость манипулирования генами диктуется конкретными задачами фундаментальных и прикладных исследований. Для понимания молекулярных механизмов функционирования отдельных генов и взаимосвязанных генетических систем большое значение имеет работа с изолированными генами. Такие исследования позволяют определить границы генов, выделить их в чистом виде и идентифицировать элементы структуры, существенные для функционирования. Доказательством функциональной значимости выделенного участка генома может быть только его нормальная экспрессия в модельной генетической системе. Поэтому следующим этапом исследования выделейного гена всегда является перемещение его в такую генетическую систему, где экспрессия гена легко обнаруживается. Результаты экспрессии оценивают либо по появлению белкового продукта, кодируемого исследуемым геном, либо по изменению функций биологической системы вследствие появления в ней новой ферментативной или другой активности, например по компенсации присутствующей в этой системе мутации. Таким образом, в результате исследования структуры конкретного гена и моделирования его экспрессии в искусственной генетической системе можно понять особенности его функционирования в живом организме. Подобный подход может быть успешно применен как к известным генам, которые выделяются целенаправленно, так и к неидентифицированным ранее последовательностям нуклеотидов, функциональную значимость которых определяют лишь после выделения их в чистом виде. Последний подход реализуется в так называемой обратной генетике.
В настоящее время с помощью методов генной инженерии получены данные о структуре и функционировании генов разнообразных организмов, что дало возможность перейти на качественно новый уровень генетических исследований. Это, во-первых, возможность переноса генова в ное для него генетическое окружение с дальнейшей его экспрессией, что ведет к изменению свойств организма, в геном которого вводится ген (например создание продуцентов биологически активных веществ или трансгенных животных), а также осуществление генотерапии наследственных и приобретенных заболеваний путем искусственного замещения мутантных аллелей. Во-вторых, стало реальным конструирование новых генов путем объединения in vitro как известных, так и новых, искусственно синтезированных последовательностей нуклеотидов. Этот подход используется в белковой инженерии для исследования функциональной значимости отдельных аминокислот и доменов в полипептидных цепях ферментов, а также для создания новых белков. В- третьих, в современной биотехнологии появилась возможность применять изолированные гены в составе генно-инженерных конструкций для получения пищевых продуктов и биологически активных веществ белковой природы
1.3.3 Предмет рекомбинантной ДНК-биотехнологии
Практическое использование рекомбинантных ДНК различного происхождения составляет основу рекомбинантной ДНК-биотехнологии, или сокращенно рДНК-биотехнологии.
Сущность рДНК-биотехнологии заключается в том, что нуклеотидная последовательность, которую необходимо выделить или размножить, ковалентно встраивается в самореплицируюшиеся молекулы нуклеиновой кислоты, называемые векторами. Далее такая последовательность нуклеотидов в составе вектора вводится в клетки про- или эукариотического организма, и эти гибридные клетки в селективных условиях, обеспечивающих сохранение вектора внутри клеток, выращивают на питательной среде. В результате образуется клон клеток, теоретически содержащих идентичные векторные молекулы с одной и той же вставкой чужеродной последовательности нуклеотидов. Поскольку объединение молекул клонируемой последовательности нуклеотидов и вектора является не чем иным, как рекомбинацией in vitro, такие гибридные молекулы называют рекомбинантными молекулами. В настоящее время разработаны многочисленные методы, позволяющие выделять определенные последовательности нуклеотидов из сложной смеси фрагментов хромосомной ДНК, а также осуществлять обмен между строго определенными фрагментами генов и другими последовательностями нуклеиновых кислот. Во всех этих реакциях, как правило, используются высокоочищенные препараты нуклеиновых кислот и ферментов нуклеинового обмена /8/.
рДНК-биотехнологию подразделяют на следующие этапы: получение чужеродной ДНК, разрезание полученной ДНК на фрагменты и их очистка,
включение фрагмента чужеродной ДНК в векторную плазмиду и получение рекомбинантной ДНК, или р-ДНК, введение р-ДНК в клетки-реципиенты и клонирование генов, амплификация и экспрессия р-ДНК.
1.3.4 Получение чужеродной ДНК
Чужеродную ДНК можно получить с помощью химического или ферментативного синтеза, либо из любого организма с последующим определением её первичной структуры.
Синтетически получают гены, в случае известной аминокислотной последовательности белка. Так синтезированы гены, кодирующие образование гормонов: про-инсулин человека, интерферон а и а2.
Изоляция структурного гена из генома клетки возможна только при работе с прокариотами, гены которых не содержат интронов. У эукариот ген состоит не только из участков, где записана структура белка (экзонов), но и участков, которые не кодируют белок (интронов) /9/.
Транскрипция у эукариот происходит в ядре, при этом образуется матричная РНК (м-РНК). При её перемещении из ядра через ядерную мембрану в цитоплазму происходит созревание м-РНК - процессинг, в результате чего интроны вырезаются с помощью специальных ферментов, а экзоны сшиваются (сплайсинг). При работе с эукариотами используют синтез структурного гена путем обратной транскрипции, то есть получение ДНК путём копирования м- РНК. Для получения комплементарной ДНК в качестве матрицы используют зрелую м-РНК, не содержащую интронов
1.3.5 Конструирование рекомбинантных ДНК
Основными инструментами для молекулярного конструирования являются два типа ферментов:
а) рестриктирующие эндонуклеазы (рестриктазы), необходимые для получения фрагментов ДНК;
б) лигазы, служащие для сшивания (соединения) участков ДНК.
1.3.5.1 ДНК-рестриктазы и ДНК-метилазы
Это ферменты, впервые открытые как часть системы рестрикции- модификации ДНК у бактерий, специфически гидролизуют молекулы двухцепочечных ДНК при наличии в них определенных последовательностей нуклеотидов, называемых сайтами рестрикции.
По механизму действия и молекулярной структуре различают три типа рестриктаз. Ферменты рестрикции типа I представляют собой сложные мультимерные комплексы, построенные из трех субъединиц с молекулярной массой до 300 кДа, которые обладают рестриктазной, ДНК-метилазной и АТРазной активностями. Рестриктазы типа I для проявления своей активности требуют присутствия АТР, S-аденозилметионина и ионов Mg , они не распознают специфические последовательности нуклеотидов и в силу этого не находят широкого применения в генной инженерии. Рестриктазы типа II узнают специфические последовательности нуклеотидов в точке расщепления ДНК или непосредственной близости от нее, требуют для проявления активности наличия в реакционной смеси АТР и ионов Mg2" и чаще всего используются при молекулярном клонировании. Ферменты типа III объединяют все прочие рестрикционные эндонуклеазы. Они также активны только в присутствии АТР и ионов Mg2 и не проявляют абсолютной зависимости от S-аденозилметионина. Названия рестриктаз складываются из первой буквы родового и двух букв видового названия бактерий, в которых они обнаружены, например Есо - Е. coli. В том случае, когда различные по специфичности действия рестриктазы присутствуют в клетках разных штаммов одного вида бактерий, в название рестриктазы вводят дополнительную букву, например рестриктазы Hinc и Hind выделены из бактериальных клеток Haemophilus influenzae, штаммы с и d. Цифры, следующие за буквенными обозначениями, отражают последовательность открытия соответствующих рестриктаз в клетках бактерий одного вида, например Hael, Haell и НаеШ из Н. aegipticus. Рестриктазы типа II - основной инструмент генной инженерии. Большинство рестриктаз типа II специфически узнают на ДНК тетра- и гексануклеотидные последовательности, а по крайней мере три из них - октануклеотиды. Чем короче олигонуклеотидная последовательность сайта рестрикции, узнаваемого рестриктазой, тем чаще он встречается в случайной последовательности нуклеотидов, в которой каждый из четырех нуклеотидов представлен с одинаковой частотой (50% А-Т-пар и 50% G-C-nap). Так, случайная тетрануклеотидная последовательность встречается в среднем через каждые 256 п.о., а гексануклеотидная - через каждые 4096 п.о. Однако в природных ДНК распределение нуклеотидов может заметно отличаться от случайного. Например, для эукариотических ДНК характерна низкая частота встречаемости динуклеотида CpG и соответственно сайтов рестрикции, содержащих эти динуклеотиды (рестриктазы Hhal, Hpall, TaqI, Thai, Aval, Haell, Hindll, Sail, Smal, Xhol, Xmal). Существенное отклонение частоты встречаемости сайтов рестрикции от ожидаемого при случайном их распределении вдоль ДНК свойственно и хромосомам термофильных бактерий, которым, напротив, свойственно (хотя и не во всех случаях) обогащение по G-C-парам. Для большинства сайтов, узнаваемых рестриктазами типа II, характерно наличие в них симметрии второго порядка, т.е. узнаваемые ими последовательности представляют собой палиндромы, например у рестриктазы EcoRI - 5'-GAATTC- 3'. Это означает, что нуклеотиды, расположенные в каждой из цепей на равном расстоянии от оси симметрии, комплементарны друг другу. Если точки расщепления противоположных цепей ДНК смещены друг относительно друга в сайте рестрикции, то образующиеся в результате рестрикции концы ДНК содержат выступающие одноцепочечные участки. Поскольку такие участки комплементарны сами себе и друг другу и могут между собой взаимодействовать, их часто называют "липкими" концами. В "липких" концах выступающим одноцепочечным участком может быть как 5'-, так и З'-конец Формальным признаком образования 5'- или 3'-выступающих "липких" концов в сайтах рестрикции является расположение точки расщепления цепей ДНК в последовательности, используемой для обозначения сайта рестрикции, слева или справа от оси симметрии соответственно. У некоторых рестриктаз точки расщепления обеих цепей ДНК расположены непосредственно друг под другом в сайте рестрикции. В этом случае после расщепления ДНК "липких" концов не образуется, а получаются так называемые "тупые" концы, в которых нет выступающих одноцепочечных участков ДНК. Имеется одно принципиальное функциональное различие между 5'- и 3'-выступающими "липкими" концами - последние невозможно пометить путем их достройки ДНК-полимеразой. Эту особенность следует иметь в виду при выборе рестриктаз для получения рестрикционных фрагментов ДНК, которые предполагается использовать в качестве зондов.
Рестриктазы являются высокоспецифическими ферментами. Однако для поддержания этой специфичности in vitro необходимо соблюдать в реакционной смеси оптимальные условия для действия ферментов. При нарушении таких условий у некоторых рестриктаз начинает проявляться вторичная (так называемая штриховая) активность. Так, рестриктаза EcoRI расщепляет последовательность GAATTC при рН 7,3, 100 мМ NaCl в присутствии 5 мМ MgC12, однако при изменении значений рН, понижении концентрации NaCl или замене ионов Mg2+ на Мп2+, а также в присутствии органических растворителей у фермента появляется тенденция к расщеплению более короткой последовательности ААТТ (так называемая активность EcoRI). К рестриктазам, обладающим подобными свойствами, относятся также BamHI, BstI, Bsul, Ddel, Hhal, PstI, Sail, SstI, Xbal.
В природе существуют антагонисты рестриктаз - метилазы. Это ферменты, которые катализируют реакции метилирования А или Ц в немногих специфических сайтах в хромосомах, в результате чего метилированная ДНК оказывается нечувствительной к атакам рестриктазами. Метилазы используются для ограничения числа сайтов рестрикции и получения более крупных фрагментов ДНК с помощью рестриктаз.
Большинство штаммов Е. coli содержит два типа ферментов, метилирующих ДНК: dam- и dcm-метилазы. Первая осуществляет перенос метальных групп в N-положение аденина в последовательности GATC. В таком случае многие рестриктазы (например Bell, Mbol или Clal), в состав сайтов
рестрикции которых входит данная метилированная последовательность, перестают расщеплять ДНК по этим сайтам. Аналогичное действие на некоторые рестриктазы, например EcoRII, оказывает и dcm-метилаза, осуществляющая метилирование остатков цитозина по положению С5 в последовательностях CMeCAGG и CMeCTGG.
1.3.5.2 ДНК-лигазы
Создание фосфодиэфирных связей в одноцепочечных разрывах двухцепочечной ДНК с помощью ДНК-лигаз является, наряду с рестрикцией, одним из важнейших этапов получения рекомбинантных ДНК in vitro. Наибольшее применение в генно-инженерных исследованиях находит ДНК-лигаза бактериофага Т4. Реакция лигирования протекает в два этапа. Вначале образуется промежуточный комплекс фермент-АМР (этап 1), после чего остаток AMP переносится на 5'-фосфатную группу концевого нуклеотида в точке разрыва ДНК (этап 2). Образовавшаяся фосфодиэфирная связь гидролизуется во время нуклеофильной атаки З'-ОН группы соседнего нуклеотида, что приводит к образованию новой фосфодиэфирной связи, восстанавливающей целостность сахаро-фосфатного остова ДНК. Т4-ДНК- лигаза осуществляет соединение фрагментов двухцепочечной ДНК, обладающих комплементарными "липкими" или "тупыми" концами. Как следует из механизма реакции, необходимым условием протекания лигирования является наличие 5'-концевого фосфата и З'-концевого гидроксила в точках разрыва цепей ДНК. При этом эффективность соединения фрагментов ДНК по "тупым" концам Т4-ДНК-лигазой возрастает в присутствии Т4-РНК-лигазы, которая осуществляет ковалентное соединение 5'-фосфорилированных концов одноцепочечных ДНК или РНК с З'-ОН группами одноцепочечных нуклеиновых кислот.
1.3.6 Векторы
Ферменты, описанные в предыдущем разделе, позволяют производить тонкие манипуляции как с протяженными молекулами ДНК, так и с их фрагментами. В частности, с помощью рестриктаз можно с большой точностью разрезать молекулы ДНК, а образовавшиеся в результате фрагменты соединять в любой желаемой последовательности друг с другом, восстанавливая сахаро- фосфатный остов молекулы ДНК с помощью ДНК-лигазы. Однако с использованием только этих ферментов еще нельзя решить одну из основных методических задач молекулярной генетики - выделение любой требуемой нуклеотидной последовательности в чистом виде и в количестве, достаточном для исследования этих последовательностей биохимическими методами. Исключение составляет метод ПЦР, однако его применение ограничивается короткими последовательностями нуклеотидов.
Основная идея, позволяющая решать эту задачу, заключается в том, чтобы присоединить исследуемые фрагменты ДНК к молекуле-переносчику, которая могла бы автономно существовать внутри бактериальных или эукариотических клеток в виде одной или нескольких копий и передаваться вместе со встроенным в нее фрагментом ДНК от одной клетки к другой. Такие молекулы- переносчики фрагментов нуклеиновых кислот были созданы, их называют векторами, и они являются одним из важнейших инструментов генной инженерии.
Идеальная векторная молекула должна обладать несколькими обязательными свойствами. Во-первых, любой вектор должен длительное время существовать в популяции клеток-хозяев, т.е. реплицироваться автономно или вместе с хромосомами клеток. Во-вторых, в любом векторе должны быть биохимические или генетические маркеры, которые позволяли бы обнаруживать его присутствие в клетках. В-третьих, структура векторной молекулы должна допускать встраивание в нее чужеродной последовательности нуклеотидов без нарушения ее функциональной целостности. Для конструирования векторов в генной инженерии используют небольшие молекулы нуклеиновых кислот, способные к автономной репликации в бактериальных и эукариотических клетках - плазмиды, хромосомы вирусов, а также фрагменты хромосом эукариотических клеток /15, 16/.
Различные векторы для клонирования ДНК и их рестрикционные карты приведены на рисунке 1.4

Первые эффективные векторы для клонирования фрагментов чужеродной ДНК, не утратившие своего значения и поныне, были получены с использованием бактериальных плазмид. Большая серия векторных плазмид, обозначенных символом pBR, создана на основе репликона природной плазмиды ColEI, придающей клеткам Е. coli устойчивость к колицину путем его объединения с генами устойчивости к антибиотикам. Таким образом, бактериальные клетки, несущие подобные комбинированные плазмиды, приобретали устойчивость к соответствующим антибиотикам, и их было легко отличить от бесплазмидных клеток путем простого посева на питательную среду с антибиотиками. Генетическая карта одного широко распространенного вектора этой серии - pBR322 изображена на рис. II.5,а.
Такая плазмида представляет собой кольцевую ковалентно замкнутую молекулу ДНК длиной 4363 и.о. Последовательность нуклеотидов pBR322 полностью известна. Плазмида содержит гены устойчивости к тетрациклину (Тс) и ампициллину (Ар), которые были перенесены в плазмиду pBR322 из плазмиды pSClOl и транспозона ТпЗ соответственно. Оба этих гена являются селектируемыми генетическими маркерами плазмиды, т.е. позволяют проводить отбор бактериальных клеток с плазмидой pBR322 по их способности к росту на питательных средах в присутствии тетрациклина и(или) ампициллина. Плазмида pBR322 содержит также обеспечивающий ее стабильную репликацию в клетках Е. coli участок ДНК, который включает область начала репликации (ori). Характерной чертой плазмиды pBR322, как и любого современного вектора, является наличие в ней нескольких уникальных сайтов рестрикции, обозначенных на генетической карте. Следует иметь в виду, что встраивание в плазмиду клонируемых чужеродных фрагментов ДНК по сайтам рестрикции, расположенным в генах Ар или Тс (например PstI или BamHI), будет нарушать целостность этих генов и их функциональную активность. В результате происходит утрата бактериальными клетками, содержащими рекомбинантные плазмиды, устойчивости к соответствующим антибиотикам.
1.3.7 Введение вектора в клетку
Клетка-реципиент—эта та среда, в которой может функционировать рекомбинантная ДНК в виде вектора. Такие клетки называют пермиссивными. К ним предъявляют следующие требования:
· не разрушаются чужеродной ДНК или РНК собственными ферментам;
· срабатывает механизм репликации вектора;
· проявляется выраженная активность промотера и/или терминатора транскрипции р-ДНК;
· отмечается полный сплайсиг м-РНК ;
· наблюдается эффективная трансляция м-РНК;
· нет выраженной активности пептидогидролаз, катализирующих реакций гидролиза экспрессируемых чужеродных белков.
Часто в качестве пермиссивных клеток используют Е. coli, Bacillus subtilis, В. stearothermophilus, В. brevis, Saccharomyces cerevisiae.
Для включения вектора в пермессивные клетки используют различные методы: трансформация, инфекция, микроинъекция.
Клетки, способные адсорбировать и поглощать вектор, называют компетентными. У таких клеток изменены свойства клеточной стенки: снижен поверхностный заряд и повышена чувствительность к осмотическому шоку.
Компетентность клеток можно повысить, используя соли кальциялибо совместно соли кальция, магния, рубидия. Кроме того, применяют глубокое замораживание и оттаивание, а также электропорацию (кратковременное воздействие электрического поля высокой напряженности, которое вызывает разрушение цитоплазматической мембраны с образованием пор).
1.3.8 Обнаружение рекомбинантного клона
Обнаружение рекомбинантных микроорганизмов проводят, используя встроенные в вектор маркеры (например, резистентности к какому-либо антибиотику).
Подробнее рассмотрим обнаружение рекомбинантного клона на примере рассмотренной выше плазмиды pBR322. В данном случае легко различить бактериальные клетки, не содержащие плазмиды (не растут в присутствии ампициллина и тетрациклина), клетки с плазмидой, не содержащей вставки клонируемой ДНК (растут в присутствии обоих антибиотиков), и клетки с рекомбинантными плазмидами (в зависимости от локализации вставки могут расти на среде только с одним из двух вышеупомянутых антибиотиков). Следовательно, наличие в векторных молекулах селектируемых маркеров резко повышает эффективность клонирования из-за возможности проведения быстрого отбора рекомбинантных плазмид на селективных питательных средах.
Помимо генов устойчивости к антибиотикам в качестве селектируемых маркеров используют и другие гены или их фрагменты. В частности, для этих целей часто применяются гены различных ферментов, присутствие которых в клетках в составе плазмиды обнаруживают по появлению соответствующей ферментативной активности.
2.Научно-исследовательская часть
2.1 Характеристика объекта исследования
Целью выполненной нами работы явилось получение Ad-вектора на основе генома аденовируса птиц CELO (chicken embryo lethal orphan или fowl adenovirus type 1) методом гомологичной рекомбинации в Escherichia coli ( E. coli, кишечной палочке).
Преимущества плазмиды, рекомбинирующей в Е. coli, перед плазмидами, рекомбинирующими в культуре клеток, следующие: легкость выращивания бактериальных клеток по сравнению с трудоемким процессом ведения культур перевиваемых или полуперевиваемых эукариотических клеток; менее жесткие условия стерильности при выращивании бактерий; дешевизна посевного бактериального материала.
В настоящее время аденовирусы (Ad) хорошо изучены в качестве этиологических агентов различных инфекционных заболеваний человека и животных как модели для многочисленных исследований в молекулярной биологии, включая изучение сплайсинга мРНК, репликации ДНК, транскрипции и клеточной трансформации. Ad широко применяют в качестве векторов для транзиентной экспрессии генов в клетках млекопитающих. Известно большое количество методов внесения изменений в геном Ad для создания векторов, способных трансдуцировать клетки млекопитающих in vivo, исследуется применение векторов на основе Ad человека в генной терапии. Векторы на основе генома Ad человека (Ad2, Ad5 и другие) имеют ряд недостатков: предсуществующий иммунный ответ организма человека на данные Ad, ограниченный спектр клеток, в которые Ad человека эффективно проникают; наращивание препаративных количеств Ad человека в культуре клеток является трудоемким и дорогостоящим процессом. В связи с этим вызывают интерес исследования, направленные на создание векторных систем на основе генома Ad животных, в первую очередь птиц, которые были бы способны проникать в различные типы клеток с большой эффективностью, на которые не было бы предсуществующего иммунного ответа в организме человека и наращивание которых в куриных эмбрионах было более технологичным и экономически выгодным.
По общей структуре вирус CELO сходен с вирусами млекопитающих, однако размер геномной ДНК у вируса CELO (44 тысячи пар оснований-т.н.п.) больше, чем у Ad5 (36 т.н.п.). Особенностью структуры капсида вириона CELO является наличие двух фиберов различной длины, отходящих от одного основания пентона, в отличие от большинства других Ad млекопитающих и птиц, имеющих один фибер. Известно, что CELO не способен к полноценному циклу репликации в клетках человека или млекопитающих и является по отношению к данным хозяевам дефектным. Была показана возможность получения Ad-вектора на основе генома CELO.
Преимуществами векторов на основе генома CELO являются отсутствие первичного иммунного ответа в организме человека и животных, большая пакующая емкость и более высокая физическая стабильность вирионов, чем Ad человека. Наращивание вируса CELO возможно в куриных эмбрионах в
больших количествах (10 вирусных частиц из одного эмбриона), что значительно снижает стоимость получения препаративных количеств вируса.
2.2 Описание эксперимента
Для получения рекомбинантного аденовируса CELO методом котрансформации с ДНК вируса CELO дикого типа сконструированная плазмида должна содержать правый и левый фрагменты ДНК указанного вируса. Мы использовали плазмиду pCBEl, содержащую правый концевой фрагмент генома вируса CELO от 88.8 доЮО единиц карты (ед. к.). Участок генома от 95,3 до 99,7 ед. к., который не является существенным для репликации вируса, был делетирован из pCBEl по сайтам рестриктазы EcoRV. Указанным способом была получена плазмида pCBEARV. Полученная плазмида была подвергнута гидролизу рестриктазой Hind3 с последующим дефосфорилированием, а затем обработке олигонуклеотидом , содержащим сайт распознавания рестриктазы Рас 1.
Левый фрагмент вируса CELO был клонирован в плазмиду pGEM-T путем постановки полимеразной цепной реакции (ПЦР). В качестве праймеров были использованы олигонуклеотиды, содержащие следующие сайты рестрикции: первый - Bst и Pad, второй—Asc, Sfi и Есо721.Число циклов амплификации составило 15.
Следующий этап заключался в проведении гидролиза полученных плазмид; PacpCBEARV по сайту рестрикции Smal, CELOleftpGEM-T путем обработки рестриктазами Bstl 1071 и Есо721. Из полученных фрагментов была сконструирована плазмида, несущая правый и левый концы генома аденовируса CELO. Недостатком полученной плазмиды являлось наличие высококопийного ori ( ori—участок, в котором начинает расплетаться ДНК при помощи ДНК- полимеразы). Учитывая наличие двух сайтов рестрикции по Pad, ori было сменено при помощи рестрикции по указанным сайтам и последующим соединением липких концов образованного фрагмента с частью плазмиды pTG3601, гидролизованной также по двум сайтам Pad.
Полученная плазмида была рестриктирована по Ascl и котрансформирована с геномом вируса CELO по гомологичным участкам. Таким образом нами был получен Ad-вектор для экспрессии генов на основе генома аденовируса птиц CELO. Указанная схема представлена на рисунке 2.1.
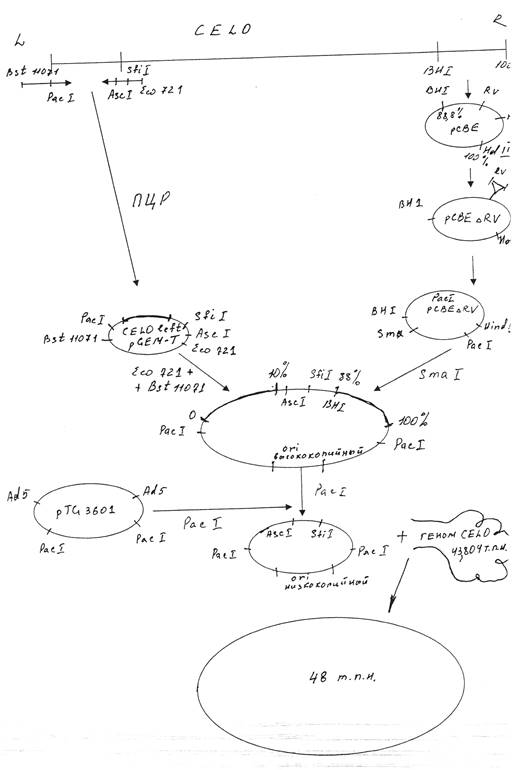
2
2.3 Обработка экспериментальных данных
Плазмида с измененным ori была подвергнута наращиванию в Е. coli препаративных количествах. Для доказательства идентичности полученных плазмид с исходной был проведен рестрикционный анализ. В лунки 0.8% агарозного геля были внесены следующие пробы: в первую—маркер молекулярного веса (геном фага X, разрезанный рестриктазой РСТ, см. рисунок 2, б));
во вторую — плазмида, нарощенная в препаративных количествах; в третью - первоначально полученную плазмиду. По результату электрофореза, представленному на рисунке 2.2, можно судить об идентичности молекулярного состава указанных плазмид.

Плазмида, полученная после рекомбинации с геномом CELO и имеющая 48 тысяч пар оснований, была подвергнута обработке рестриктазой HindHI. Указанная рестриктаза активна при следующей комбинации нуклеотидов:
![]()
Количество сайтов рестрикции в полученной плазмиде и их местоположение указаны на рисунке:

Пробы после рестрикции были подвергнуты электрофорезу. В первую, вторую и третью лунку помещены целевые плазмиды, полученные из разных клонов Е. coli. В четвертую лунку помещена плазмида, неподвергавшаяся рестрикции, в пятую — маркер молекулярного веса.
Результаты рестрикционного анализа свидетельствуют об идентичности полученных фрагментов с ожидаемыми.
Заключение
В заключении данной работы рассмотрим наиболее существенные и практически значимые достижения рекомбинантной ДНК-биотехнологии, а также перспективы её развития.
Начиная с момента открытия двойной спиральной структуры а особенно после расшифровки генома человека, научные изыскания приносят значительные практические результаты, наиболее перспективные из которых следующие: конструирование рекомбинантных организмов-продуцентов жизненно необходимых лекарственных средств, получение которых альтернативными методами невозможно или сопряжено со значительными трудностями; разработка генетических вакцин; развитие генной терапии; применение трансгенных растений и животных для интенсификации сельского хозяйства.
Сравнение структур генов секвенированных к настоящему времени геномов человека, дрозофилы нематоды, дрожжей и бактерий приводит к выводу, что все живые существа действительно произошли от общего предшественника, так как родственные гены легко идентифицируются в геномах представителей всех трех царств живых организмов. Указанный факт теоретически обосновывает возможность встраивания и успешного функционирования чужеродных генов в организме. В настоящее время широкий спектр биологически активных веществ производится рекомбинантными организмами (соматотропин, инсулин, интерфероны, интерлейкин - 2).
Одним из самых современных и перспективных направлений биотехнологии является разработка генетических вакцин. Преимуществом генетических вакцин является возможность фокусирования иммунного ответа на одном или нескольких определенных антигенах возбудителя, что принципиально невозможно при использовании традиционных вакцин и не происходит при течении инфекционного процесса, а также возможность их использования не только в превентивных, но и в терапевтических целях для лечения аутоиммунных заболеваний, аллергий и злокачественных новообразований.
К терапии нового поколения относится генная или клеточная терапия основанная на превращении стволовых клеток в любые другие необходимые для организма клетки. В настоящее время указанная выше методика находится в стадии разработки, но имеет реальную перспективу.
Литература
1. Альбертс Б., Брей Д., Льюис Дж. и др. Молекулярная биология клетки в 3-х томах. — М.: Мир, 1994. — 1558 с. — ISBN 5-03-001986-3.
2. Докинз Р. Эгоистичный ген. — М.: Мир.[82]
3. История биологии с начала XX века до наших дней. — М.: Наука, 1975. — 660 с.
4. Льюин Б. Гены. — М.: Мир, 1987. — 1064 с.
5. Пташне М. Переключение генов. Регуляция генной активности и фаг лямбда. — М.: Мир, 1989. — 160 с.[83]
6. Уотсон Дж. Д. Двойная спираль: воспоминания об открытии структуры ДНК. — М.: Мир, 1969. — 152 с.
7. Burgers P, Koonin E, Bruford E et al. (2001). «Eukaryotic DNA polymerases: proposal for a revised nomenclature». J. Biol. Chem. 276 (47): 43487-90. PMID 11579108
8. Авторозов Е.К. Лекции по микробиологии и биотехнологии – М.: Эксмо, 2000
9. Елинов М.П. Основы биотехнологии – М.: Москва, 2004
10. Егорова Н.С. Биотехнология. Учебное пособие для ВУЗов – М.: Просвещение, 2009.
Похожие рефераты:
Литература - Другое (книга по генетике)
Продукты рекомбинации: характеристика и манипулирование
рДНК-биотехнология. Способы биотрансформации клеток
Возникновение злокачественных опухолей
Изучение вопросов биотехнологии в курсе химии средней школы
Конструирование, клонирование и отбор рекомбинантных молекул ДНК
Наследственность, представления о генетическом коде, гены индивидуальности