| Скачать .docx | Скачать .pdf |
Курсовая работа: Общая характеристика нервной ткани
1. Общая характеристика нервная ткани
2.1 Классификация и функции нервных клеток
5.1 Безмиелиновые нервные волокна
5.2 Миелиновые нервные волокна
8. Понятия о рефлекторных дугах
Введение
Для согласованной деятельности различных частей такой сложной системы, как организм, необходима координационная система соответствующей сложности. Нервная система, которая интегрирует деятельность всех частей тела, является, безусловно, самой сложной из всех систем организма. Нервная ткань, из которой построена нервная система, способна к восприятию информации и обеспечивает реакцию на нее всего организма.
Восприятие информации осуществляется особыми образованиями нервной ткани, которые называются рецепторами. В зависимости от локализации раздражений различают экстерорецепторы и интерорецепторы: первые реагируют на стимулы, исходящие из окружающей организм внешней среды, вторые - на изменения, происходящие в самом организме. Кроме того, рецепторы подразделяют соответственно тем формам энергии, к которым они особо чувствительны, а именно: терморецепторы, механорецепторы, фоторецепторы, хеморецепторы, электрорецепторы.
В основе функции нервной системы лежит процесс отражения, или рефлекс (reflexus - обращение назад), основанный на отражении объективных явлений внешней или внутренней среды организма. На этой основе возникла рефлекторная теория, материалистически объясняющая принципы работы мозга. Основными функциями нервной системы является отражение явлений внешнего мира и внутренней среды организма, генерация и проведение нервных импульсов, а также интеграция деятельности всех систем, организма.
Координационная деятельность нервной системы, эндокринная регуляция и регуляторные механизмы внутриклеточных ферментных систем являются факторами, которые обеспечивают гомеостаз (homoios - одинаковый, подобный, stasis - неподвижность, стояние) организма, т.е. поддерживают постоянство его внутренней среды. Обеспечивая приспособляемость организма к условиям существования, нервная система сама обнаруживает способность к приспособлению, образуя новые рефлекторные связи. У позвоночных животных и человека нервная ткань составляет основу нервной системы, которая представлена головным и спинным мозгом, нервами, нервными узлами, или ганглиями, и нервными окончаниями.
1. Общая характеристика нервная ткани
Нервной ткани входят два вида клеток: нервные клетки, или нейроны (нейроциты), и глиальные клетки, или глиоциты. Первым присуща функция возбуждения и проведения нёрвного импульса, а вторым - опорная, трофическая, изоляционная и защитная функция. Совокупность глиоцитов составляет нейроглию. Клетки нейроглии подразделяются на две группы: глиоциты, которые относятся к макроглии, и глиальные макрофаги, которые относится; к микроглии. В свою очередь клетки макроглии подразделяются на эпендимоциты, астроциты и олигодендроциты, которые соответственно образуют эпендиму, астроглию и олигодендроглию.

2. Нервные клетки
2.1 Классификация и функции нервных клеток
Нервные клетки, или нейроны (нейроциты) (neurocytus, neuronum), различных отделов нервной системы значительно отличаются друг от друга по функциональному значению и морфологическим особенностям. Нейроны, возникающие из невральной эктодермы (нервная трубка), наследуют от ее клеток гетерополярность, типичную для эпителиальной ткани. Гетерополярность определяет дифференцировку нейронов и их форму. В зависимости от выполняемой функции нейроны делят на чувствительные (рецепторные или эффекторные), ассоциативные и эффекторные (двигательные или моторные). Первые генерируют нервный импульс под влиянием различных воздействий внешней или внутренней среды организма. Эффекторные нейроны передают возбуждение на ткани рабочих органов, побуждая их к действию. Ассоциативные (вставочные) нервные клетки осуществляют различные связи между нейронами. Нервные клетки имеют различную величину. Так, например, диаметр тела клеток зернистого слоя мозжечка колеблется от 4 до 6 мк, а размеры тела гигантских пирамидных клеток коры больших полушарий головного мозга (клетки Беца) достигают 130 мк. Столь же разнообразна и специфична для различных отделов нервной системы форма нейронов. Характерной чертой для всех зрелых нервных клеток является наличие у них отростков. Эти отростки обеспечивают проведение нервного импульса по телу человека из одной его части в другую, подчас весьма удаленную, и поэтому длина их колеблется в больших пределах - от нескольких микронов до 1-1,5 м.
–
Рис. 1. Нейроны: А – униполярный; Б – биполярный; В мультиполярный; 1 – аксон; 2 – дендрит

Рис. 2. Псевдоуниполярные нейроны. Чувствительные нейроны из полулунного узла.
По функциональному значению отростки нервных клеток делятся на два вида. Одни из них выполняют функцию отведения нервного импульса от тел нейронов и называются нейритами, или аксонами (лат. axis-ось). В процессе дифференцировки нейробласта аксон образуется раньше других отростков клетки. Все зрелые нейроны имеют один аксон. Он заканчивается концевым аппаратом или на другом нейроне, пли на тканях рабочего органа - на мышцах или железах. Второй вид отростков нервных клеток носит название дендритов (греч. dendron - дерево). Дендриты образуются в процессе дифференцировки нервных клеток позднее аксонов. Количество их у нейронов разных отделов нервной системы различно, так же как различна и их форма. В большинстве случаев они сильно ветвятся, чем и определяется их название. Дендриты воспринимают нервное раздражение и проводят нервный импульс к телу нейрона. В ряде случаев дендриты имеют на своем периферическом конце специфически устроенные воспринимающие аппараты - чувствительные нервные окончания (рецепторы).
По количеству отростков нервные клетки делятся на три группы:
униполярные клетки с одним отростком,
биполярные - клетки с двумя отростками,
мультиполярные - клетки, имеющие три и больше отростков (см. рис.1).
Наиболее распространенная форма нейронов у млекопитающих животных и человека - мультиполярные нейроны. Из многих отростков такого нейрона один представлен аксоном, тогда как все остальные являются дендритами. Биполярные клетки имеют два отростка - аксон и дендрит. Истинные биполярные клетки в теле человека встречаются редко. Такую форму имеют часть клеток сетчатки глаза, спирального ганглия внутреннего уха и некоторые другие. Однако по существу своего строения к биполярным клеткам должна быть отнесена большая группа чувствительных - так называемых псевдоуниполярных нейронов краниальных и спинальных нервных узлов (рис. 2). Эти клетки также имеют дендрит, идущий на периферию, заканчивающийся там чувствительными окончаниями (рецепторами), и аксон, несущий нервный импульс от тела клетки в центральную нервную систему. Псевдоуниполярными они называются потому, что аксон и дендрит этих клеток начинаются от общего выроста тела, создающего впечатление одного отростка, с последующим Т-образным делением его. Истинных униполярных клеток, т.е. клеток с одним отростком - аксон, в теле человека нет, если не считать униполярной формы нейробластов до периода образования дендритов.
2.2 Ядро нервной клетки
Нервные клетки человека в подавляющем большинстве содержат одно ядро. Двухъядерные нейроны и тем более многоядерные встречаются крайне редко. Исключение составляют нервные клетки некоторых ганглиев вегетативной нервной системы, а именно - сплетения предстательной железы и узлов шейки матки. В этих нервных образованиях можно иногда наблюдать нейроны, содержащие до 15 ядер.
Форма ядра нервных клеток округлая. В ядрах содержится мало хроматина, что часто придает им на окрашенных препаратах пузырькообразный вид. Располагаются ядра обычно в центре тела нейрона, реже эксцентрично. Изучение ядер нервных клеток под электронным микроскопом показало, что они отграничены от цитоплазмы клетки двумя мембранами, расположенными друг от друга на расстоянии 200 Ǻ и имеющими поры. В ядре нервных клеток имеется одно, а иногда 2 - 3 крупных ядрышка. Усиление функциональной активности нейронов обычно сопровождается увеличением объема и количества ядрышек. Ядра нервных клеток, особенно ядрышки, богаты РНК. Ряд авторов высказывает предположения, что в некоторых нейронах, характеризующихся высоким показателем ядерно-плазменного отношения (клетки-зерна мозжечка, ганглионарные клетки сетчатки и др.), значительная часть белков образуется в ядре, откуда поступает в цитоплазму и в отростки. ДНК ядра обычно мелко распылена, поэтому ядра крупных нейронов выглядят светлыми.
2.3 Цитоплазма нервной клетки
Цитоплазма нейронов содержит обычные для всех клеток органеллы. Пластинчатый комплекс в нервных клетках был впервые описан Гольджи в 1898 г. Наличие центросомы в настоящее время установлено в нейронах почти всех отделов нервной системы. Центросома лежит чаще всего около ядра нейрона, занимая всегда определенное положение в клетке. В нейробластах в период формирования нейрона центросома находится со стороны растущего отростка (аксона). В дифференцированных нейронах центросома лежит между дендритами и ядром. Митохондрии расположены как в теле нейрона, так и во всех его отростках. Особенно богата митохондриями цитоплазма нервных клеток в месте отхождения аксона и в концевых аппаратах отростков, в частности цитоплазма структур межнейрональных синапсов. Митохондрии в нервных клетках при рассмотрении в световом микроскопе имеют форму палочек, нитей и зерен. По субмикроскопическому строению они существенно не отличаются от митохондрий других клеток.
Цитоплазматическая сеть в дифференцированных нейронах представлена системой связанных между собой цистерн, пузырьков и канальцев. Их диаметр колеблется от 300 до 400 Ǻ, а в отдельных случаях достигает 800-2000 Ǻ. В совокупности они представляют трехмерную сеть двухконтурных мембран (альфа-цитомембран), ориентированных параллельно друг другу. Степень ориентации мембран в нейронах различных типов неодинакова. Максимально упорядоченно располагаются мембраны в нейронах спинного мозга. В целом цитоплазматическая сеть цитоплазмы нейронов - структура очень подвижная, изменяющаяся в соответствии с функциональным состоянием клетки.
Цитоплазма всех нервных клеток богата рибосомами, которые, как и в клетках других тканей, представлены гранулами диаметром 150-350 Ǻ. В нейробластах рибосомы распределяются в матриксе свободно по одиночке или образуют небольшие группы - полирибосомы. В дифференцированных нейронах значительная часть рибосом связана с поверхностью мембран цитоплазматической сети, которая соответствует эргастоплазме железистых или других клеток, продуцирующих белок.

Рис. 3. Тигроидное вещество в корешковом нейроне спинного мозга (схема): 1 – аксон; 2 – дендрит
Базофильное вещество (substantia basophila), или хроматофильное вещество, тигроидное вещество, глыбки Ниссля, - участки цитоплазмы с большим содержанием рибосом, а, следовательно, и РНК, интенсивно окрашивается основными красителями. В соответствии с этим на препаратах, обработанных основными красками, или специфически на РНК, в перикарионе нейронов и их дендритах выявляется зернистость. Она образует в совокупности нерезко отграниченные базофильные глыбки, впервые описанные Нисслем (рис.3).
Базофильное вещество никогда не содержится в аксоне и в его конусовидном основании (аксонном холмике). Морфологии базофильного вещества различных типов нейронов присущ ряд особенностей.
Так, в моторных клетках спинного мозга глыбки базофильного вещества крупные, неправильной угловатой формы; расположены они наиболее плотно вокруг ядра. Ближе к периферии тела клетки и в дендритах они обычно мельче, несколько вытянуты в длину и лежат реже. В чувствительных нейронах спинальных ганглиев глыбки имеют вид мелкой пылевидной зернистости. Базофильное вещество в клетках большинства узлов вегетативной нервной системы представлено мелкими зернами, расположенными в цитоплазме неравномерно, и образует нежную сеточку (узлы пограничного симпатического ствола, верхний шейный узел). В других ганглиях базофильное вещество состоит из грубых глыбок заполняющих все тело клетки (узлы солнечного сплетения, звездчатый узел) и ее дендриты.
Морфология базофильного вещества изменяется в зависимости от функционального состояния клетки. При увеличении интенсивности специфической деятельности нейрона базофилия глыбок возрастает. В условиях перенапряжения или каких-либо травм (перерезка отростков, отравление, кислородное голодание, неадекватное раздражение) глыбки распадаются и исчезают. Этот процесс получил название хроматолиза (тигролиза), т.е. растворения базофильного вещества. Хроматолиз в разных случаях имеет свои специфические особенности, соответствующие характеру травмы. Это позволяет по морфологическим изменениям базофильного вещества судить о состоянии нервных клеток в условиях патологии и эксперимента. Возвращение нейронов в нормальное состояние сопровождается восстановлением типичной для этих клеток картины базофильного вещества.
Глыбки базофильного вещества нейронов представляют собой участки цитоплазмы, соответствующие гранулярной цитоплазматической сети других клеток. Так как РНК принимает активное участие в синтезе белковых веществ, можно считать, что глыбки базофильного вещества являются частью цитоплазмы, активно синтезирующей белок, необходимый для осуществления специфической функции нейрона.
При дифференцировке нейронов в период эмбрионального развития по мере роста отростков объем цитоплазмы резко увеличивается (в 2000 раз и более), при этом в соответствии с интенсивностью синтеза белка содержание РНК в них постепенно увеличивается и оформляется базофильное вещество. Наиболее заметные сдвиги в синтезе белка, накоплении РНК и формировании базофильного вещества наблюдаются в определенные периоды развития зародыша, совпадающие с усилением деятельности нервной системы. Например, с 7-х суток развития зародыша курицы обнаруживаются его рефлекторные движения, так как к этому времени оформляются рефлекторные дуги. Появление движений совпадает с увеличением концентрации РНК в моторных клетках спинного мозга и в чувствительных клетках спинальных ганглиев. В последующие дни моторная активность зародыша ослабевает, что сопровождается снижением количества РНК в нервных клетках. Затем двигательная активность зародыша усиливается с 19-20-х суток. В это время соответственно резко увеличивается и концентрация РНК, а также связанного с ней основного белка в нервных клетках. Базофильное вещество приобретает характерные для зрелой нервной клетки форму и химический состав.
Кроме гранулярного вида цитоплазматической сети, для цитоплазмы нервных клеток характерно наличие гладкой цитоплазматической сети в виде узких трубочек и пузырьков. В тесной связи с базофильным веществом в ряде нервных клеток, например в двигательных клетках, находятся включения гликогена, который образует с ними временные связи (симплексы). Помимо того, в цитоплазме нервных клеток всегда имеются различные ферменты: оксидаза, пероксидаза, фосфатаза, холинэстераза и др.
Пигментные включения нервных клеток представлены двумя видами пигмента. Меланин в виде черных, грубых, различной величины зерен находится только в определенных отделах нервной системы, а именно - в нейронах черного вещества и голубого места, а также дорсального ядра блуждающего нерва. Желтый пигмент липофусцин, содержащий липоиды, в виде мелкой зернистости встречается в нервных клетках всех отделов нервной системы. Появляется он у человека преимущественно после 7 лет и количество его увеличивается к 30 годам жизни.
2.4 Нейрофибриллы
В цитоплазме фиксированных и обработанных солями серебра нервных клеток выявляется сеть тонких нитей - нейрофибриллы (рис.4). В отростках нейронов нейрофибриллы располагаются параллельно друг другу. В теле нервной клетки они ориентированы различно и в совокупности образуют густую связь. Нейрофибриллярный аппарат представляет собой морфологическое выражение правильной, линейной ориентации белковых молекул нейроплазмы. Изучение живых нефиксированных нервных клеток в культурах тканей, а также клеток, фиксированных при различных эксперементальных условиях, показало, что Нейрофибриллярный аппарат - структура весьма подвижная и при различных функциональных состояниях выражена не одинаково.

Рис. 4. Нейрофибриллярный аппарат нейрона (схема)
При электронной микроскопии в цитоплазме нервных клеток структуры, соответствующие микроскопически видимым нейрофибриллам, не обнаружено, но выявляются тонкие нити диаметром 60-100 Ǻ - нейрофиламенты и трубочки - нейротубулы диаметром 200-300 Ǻ. Очевидно они и представляют собой те комплексы белковых молекул, которые при агрегации и импрегнации азотнокислым серебром приобретают вид нейрофибрилл.
2.5 Нейросекреторные клетки
Наряду с описанными выше нейронами имеются группы нервных клеток, например нейроны некоторых ядер гипоталамической области головного мозга, обладающие секреторной деятельностью. Нейросекреторные клетки имеют ряд специфических морфологических признаков. Это крупные нейроны. Цитоплазма их бедна базофильным веществом; оно преимущественно располагается по периферии тела клеток. В цитоплазме нейронов и в аксонах находятся различной величины гранулы и капли секрета, содержащие белок, а в некоторых случаях - липоиды и полисахариды. Гранулы нейросекрета нерастворимы в воде и спирте. Многие нейросекреторные клетки имеют ядра неправильной формы, что свидетельствует об их высокой функциональной активности.
2.6 Зеркальные нейроны
В настоящее время некоторые ученые выделяют зеркальные нейроны. Они обнаружены недавно и еще не признаны другими научными сотрудниками. Зеркальные нейроны на стадии изучения. Конкретных функций и свойств этих нейронов неизвестно, но ученые предполагают, что одна из их задач - это “сканирование" информации с этих нейронов (например: другого человека), вследствие чего мы понимаем его настроение, о чем он думает и т.д., глядя на него (это простейший пример). Факт гистогенеза и регенерации зеркальных нейронов еще не известен.
3. Нейроглия ( Neuroglia)
Кроме нервных клеток, в состав нервной ткани входят многочисленные и весьма различные по функциональному значению клеточные элементы - нейроглия (греч. glia - клей). Они выполняют в нервной ткани опорную, разграничительную, трофическую, секреторную и защитную функции. Все элементы нейроглии делятся на два генетически различных вида: глиоциты (макроглия) и глиальные макрофаги (микроглия). Глиоциты развиваются одновременно с нейронами из спонгиобластов нервной трубки. Среди глиоцитов различают эпендимоциты, астроциты и олигодендроглиоциты. Глиальные макрофаги - производные мезенхимы.
4. Макроглия
Эпендимоциты ( ependymocyti) образуют плотный слой клеточных элементов, выстилающих спинномозговой канал и все желудочки мозга. В процессе гистогенеза нервной ткани эпендимоциты дифференцируются первыми из спонгиобластов нервной трубки (рис.5) и выполняют в этой стадии развития разграничительную и опорную функции. Вытянутые тела спонгиобластов на внутренней поверхности нервной трубки образуют слой эпителиоподобных клеток. На поверхности клеток, обращенной в полость канала нервной трубки, дифференцируются реснички, которые, очевидно, своим сокращением способствуют движению цереброспинальной жидкости. Базальные концы эпендимоцитов снабжены длинным отростком, который, разветвляясь, пересекает всю нервную трубку, образуя ее поддерживающий аппарат. Эти отростки эпендимоцитов, достигая внешней поверхности нервной трубки, принимают участие в образовании наружной разграничительной мембраны, отделяющей вещество трубки от других тканей. В постэмбриональном периоде развития реснички в эпендимных клетках постепенно утрачиваются и сохраняются лишь в некоторых местах центральной нервной системы, в водопроводе мозга.
Некоторые эпендимоциты выполняют секреторную функцию, выделяя различные активные вещества прямо в полость мозговых желудочков или кровь. Например, в области задней комиссуры головного мозга эпендимоциты образуют особый "субкомиссуральный орган", выделяющий секрет, возможно, участвующий в регуляции водного обмена.
Эпендимные клетки на поверхности сосудистых сплетений желудочков мозга кубической формы. У новорожденных эти клетки имеют на своей поверхности реснички. Под электронным микроскопом видно, что и позднее на апикальном полюсе клеток сохраняются своеобразные выросты. На базальном полюсе оболочка клеток образует многочисленные и глубокие складки. Цитоплазма содержит крупные митохондрии и различные включения: жир, пигмент и др.

Рис. 5. Эпендима желудочка мозга: 1 – реснички; 2 – эпендимоцит
При введении в кровяное русло животного трипановой сини последняя также накапливается в цитоплазме этих клеток. Все это подтверждает мнение о том, что эпендимные клетки сосудистых сплетений принимают активное участие в процессах образования цереброспинальной жидкости.
Астроциты ( astrocyti). Опорный аппарат центральной нервной системы представлен огромным количеством мелких клеток с многочисленными расходящимися во все стороны отростками - астроцитами. Различают два вида астроцитов: плазматические и волокнистые (Рис.6). Между ними имеются и переходные формы.
Плазматические астроциты лежат преимущественно в сером веществе центральной нервной системы. Они характеризуются наличием крупного округлого бедного хроматином ядра и большим количеством сильно разветвленных коротких отростков. Цитоплазма астроцитов богата митохондриями. Большое количество митохондрий в цитоплазме плазматических коротколучистых астроцитов говорит об их участии в обменных процессах. О том же свидетельствует и их активность в условиях патологии. Например, при дегенерации нервных элементов в цитоплазме коротколучистых астроцитов накапливаются различные продукты распада и особенно включения липоидов.
Волокнистые астроциты располагаются главным образом в белом веществе мозга. Эти клетки имеют 20-40 гладкоконтурированных, длинных, слабо ветвящихся отростков. В периферической зоне цитоплазма тел клеток и отростков продолжается в глиальные волокна, которые в совокупности образуют в виде плотной сети поддерживающий аппарат мозга.

Рис. 6. Различные виды нейроглии: А – Протоплаэматические астроциты; Б – волокнистые астроциты; В – олигодендроглия; Г – микроглия (глиальные макрофаги).
Отростки астроцитов на кровеносных сосудах и на поверхности мозга своими концевыми расширениями формируют разграничительные мембраны.
Олигодендроглиоциты (oligodendrogliocyti). Это самая многочисленная группа клеток нейроглии (см. Рис. 6). Олигодендроглиоциты окружают тела нейронов в центральной и периферической нервных системах, находятся в составе оболочек нервных волокон и в нервных окончаниях. В различных отделах нервной системы клетки олигодендроглии имеют различную форму. В сером веществе мозга они небольших размеров: от тел клеток, имеющих овальную или угловатую форму, отходит несколько коротких и слабо разветвленных отростков. Особенности строения олигодендроглиоцитов в составе периферических нервных узлов, волокон и окончаний будут описаны в соответствующих разделах.
Изучение под электронным микроскопом показало, что по плотности цитоплазмы клетки олигодендроглии приближаются к нервным и отличаются от них тем, что не содержат нейрофиламентов. Функциональное значение этих клеток очень разнообразно. Они выполняют трофическую функцию, принимая участие в обмене веществ нервных клеток. Олигодендроглиоциты играют значительную роль в образовании оболочек вокруг отростков клеток, а также в процессах дегенерации и регенерации нервных волокон. Клетки олигодендроглии в составе нервных окончаний участвуют в процессах рецепции (восприятия) и передачи нервного импульса.
Микроглия , или глиальные макрофаги (macrophagus glialis), мезоглия, клетки Гортега, являются производными мезенхимы. Основная их функция - фагоцитоз. Глиальные макрофаги небольших размеров, преимущественно отростчатой формы, способны к амебоидным движениям. Два-три более крупных отростка имеют на своей поверхности короткие вторичные и третичные ветвления. Ядра клеток вытянутой или треугольной формы, богаты хроматином (см. Рис. 6). При раздражении клеток микроглии их форма меняется, отростки втягиваются, клетки приобретают специфический характер, округляются. В таком виде они носят название зернистых шаров.
5. Нервные волокна

Рис. 7. Миелиновые нервные волокна из седалищного нерва лягушки, обработанного тетраоксидом осмия: 1 – слой миелина; 2 – соединительная ткань; 3 – нейролеммоцит; 4 – насечки миелина; 5 – перехват узла

Рис. 8. Межмышечное нервное сплетение кишечника кошки: 1 – безмиелиновые нервные волокна; 2 – ядра нейролеммоцитов
Отростки нервных клеток обычно одеты глиальными оболочками и вместе с ними называются нервными волокнами. Так как в различных отделах нервной системы оболочки нервных волокон значительно отличаются друг от друга по своему строению, то в соответствии с особенностями их строения все нервные волокна делятся на две основные группы - миелиноеые (рис.7) и безмиелиновые волокна (рис.8). Те и другие состоят из отростка нервной клетки (аксона или дендрита), который лежите в центре волокна и поэтому называется осевым цилиндром, и оболочки, образованной клетками олигодендроглии, которые здесь называются леммоцитами (шванновскими клетками).
5.1 Безмиелиновые нервные волокна
Находятся они преимущественно в составе вегетативной нервной системы. Клетки олигодендроглии оболочек безмиелиновых нервных волокон, располагаясь плотно, образуют тяжи цитоплазмы, в которых на определенном расстоянии друг от друга лежат овальные ядра. В безмиелиновых нервных волокнах внутренних органов часто в одной такой клетке располагается не один, а несколько (10-20) осевых цилиндров, принадлежащих различным нейронам. Они могут, покидая одно волокно, переходить в смежное. Такие волокна, содержащие несколько осевых цилиндров, называются волокнами кабельного типа. При электронной микроскопии безмиелиновых нервных волокон видно, что по мере погружения осевых цилиндров в тяж леммоцитов последние одевают их как муфта.
Оболочка леммоцитов при этом прогибается, плотно охватывает осевые цилиндры и, смыкаясь над ними, образует глубокие складки, на дне которых и располагаются отдельные осевые цилиндры. Сближенные в области складки участки оболочки леммоцита образуют двойную мембрану - мезаксон, на которой как бы подвешен осевой цилиндр (рис.9).
Так как оболочка леммоцитов очень тонка, то ни мезаксона, ни границ этих клеток под световым микроскопом нельзя рассмотреть, и оболочка безмиелиновых нервных волокон в этих условиях выявляется как однородный тяж цитоплазмы, одевающий осевые цилиндры. С поверхности каждое нервное волокно покрыто базальной мембраной.
![]()

Рис. 9. Схема продольного(А) и поперечного (Б) сечения безмиелиновых нервных волокон: 1 – ядро леммоцита; 2 – осевой цилиндр; 3 – митохондрии; 4 – граница леммоцитов; 5 – мезаксон.
5.2 Миелиновые нервные волокна
Миелиновые нервные волокна значительно толще безмиелиновых. Диаметр поперечного сечения их колеблется от 1 до 20 мк. Они также состоят из осевого цилиндра, одетого оболочкой из леммоцитов, но диаметр осевых цилиндров этого типа волокон значительно больше, а оболочка сложнее. В сформированном миелиновом волокне принято различать два слоя оболочки: внутренний, более толстый, - миелиновый слой (рис.10), и наружный, тонкий, состоящий из цитоплазмы леммоцитов и их ядер.
Миелиновый слой содержит в своем составе липоиды, а поэтому при обработке волокна осмиевой кислотой он интенсивно закрашивается в темно-коричневый цвет. Все волокно в этом случае представляется однородным цилиндром, в котором на определенном расстоянии друг от друга располагаются косо ориентированные светлые линии - насечки миелина (incision myelini), ил и насечки Шмидта-Лантермана. Через некоторые интервалы (от нескольких сотен микронов до нескольких миллиметров) волокно резко истончается, образуя сужения - узловые перехваты, или перехваты Ранвье. Перехваты соответствуют границе смежных леммоцитов. Отрезок волокна, заключенный между смежными перехватами, называется межузловым сегментом, а его оболочка представлена одной глиальной клеткой.
В процессе развития миелинового волокна осевой цилиндр, погружаясь в леммоцит, прогибает его оболочку, образуя глубокую складку.

Рис. 10. Схема нейрона. 1 – тело нервной клетки; 2 – осевой цилиндр; 3 – глиальная оболочка; 4 – ядро леммоцита; 5 – миелиновый слой; 6 – насечка; 7 – перехват Ранвье; 8 – нервное волокно, лишенное миелинового слоя: 9 – двигательное окончание; 10 – миелиновые нервные волокна, обработанные осмиевой кислотой.
По мере погружения осевого цилиндра оболочка леммоцита в области щели сближается и ее два листка соединяются друг с другом своей внешней поверхностью, образуя двойную мембрану - мезаксон (рис.11).
При дальнейшем развитии миелинового волокна мезаксон удлиняется и концентрически наслаивается на осевой цилиндр, вытесняя цитоплазму леммоцита и образуя вокруг осевого цилиндра плотную слоистую зону - миелиновый слой (рис.12). Так как оболочка леммоцита состоит из липидов и белков, а мезаксон представляет собой ее двойной листок, то естественно, что миелиновая оболочка, образованная его завитками, интенсивно окрашивается осмиевой кислотой. В соответствии с этим под электронным микроскопом каждый завиток мезаксона виден как слоистая структура, построенная из белков и липидов, расположение которых типично для мембранных структур клеток. Светлый слой имеет ширину около 80-120 Ǻ и соответствует липоидным слоям двух листков мезаксона. Посредине и по поверхности его видны тонкие темные линии, образованные молекулами белка.

Рис. 11. Схема развития миелинового волокна. 1 – контакт аксолеммы и оболочки леммоцита; 2 – щель; 3 – аксолемма и оболочка леммоцига; 4 – цитоплазма леммоцита; 5 – мезаксон
Шванновской оболочкой называется периферическая зона волокна, содержащая оттесненную сюда цитоплазму леммоцитов (шванновских клеток) и их ядра. Эта зона при обработке волокна осмиевой кислотой остается светлой. В области насечек между завитками мезаксона имеются значительные прослойки цитоплазмы, благодаря чему клеточные мембраны располагаются на некотором расстоянии друг от друга. Больше того, как видно на рис.188, листки мезаксона в этой области также лежат неплотно. В связи с этим при осмировании волокна эти участки не окрашиваются.
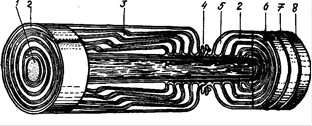
Рис. 12. Схема субмикроскопического строения миелинового нервного волокна: 1 – аксон; 2 – мезаксон; 3 – насечка миелина; 4 – узел нервного волокна; 5 – цитоплазма нейролеммоцита; 6 – ядро нейролеммоцита; 7 – нейролемма; 8 – эндоневрий
На продольном сечении вблизи перехвата видна область, в которой завитки мезаксона последовательно контактируют с осевым цилиндром. Место прикрепления самых глубоких завитков его наиболее удалено от перехвата, а все последующие завитки закономерно расположены ближе к нем у (см. рис.12). Это легко понять, если представить себе, что закручивание мезаксона идет в процессе роста осевого цилиндра и одевающих его леммоцитов. Естественно, что первые завитки мезаксона оказываются короче, чем последние. Края двух смежных леммоцитов в области перехвата образуют пальцеобразные отростки, диаметр которых равен 500 Ǻ. Длина отростков различна. Переплетаясь между собой, они образуют вокруг осевого цилиндра своеобразный воротничок и попадают на срезах то в поперечном, то в продольном направлении. В толстых волокнах, у которых область перехвата относительно коротка, толщина воротничка из отростков шванновских клеток больше, чем в тонких волокнах. Очевидно, аксон тонких волокон в перехвате более доступен для внешних воздействий. Снаружи миелиновое нервное волокно покрыто базальной мембраной, связанной с плотными тяжами коллагеновых фибрилл, ориентированных продольно и не прерывающихся в перехвате - невралеммой.
Функциональное значение оболочек миелинового нервного волокна в проведении нервного импульса в настоящее время недостаточно изучено.
Осевой цилиндр нервных волокон состоит из нейроплазмы - бесструктурной цитоплазмы нервной клетки, содержащей продольно ориентированные нейрофиламенты и нейротубулы. В нейроплазме осевого цилиндра лежат митохондрии, которых больше в непосредственной близости к перехватам и особенно много в концевых аппаратах волокна.
С поверхности осевой цилиндр покрыт мембраной - аксолеммой, обеспечивающей проведение нервного импульса. Сущность этого процесса сводится к быстрому перемещению локальной деполяризации мембраны осевого цилиндра по длине волокна. Последнее определяется проникновением в осевой цилиндр ионов натрия (Nа+ ), что меняет знак заряда внутренней поверхности мембраны на положительный. Это, в свою очередь, повышает проходимость ионов натрия в смежном участке и выход ионов калия (К+ ) на внешнюю поверхность мембраны в деполяризованном участке, в котором восстанавливается при этом исходный уровень разности потенциалов. Скорость движения волны деполяризации поверхностной мембраны осевого цилиндра определяет быстроту передачи нервного импульса. Известно, что волокна с толстым осевым цилиндром проводят раздражение быстрее тонких волокон. Скорость передачи импульса миелиновыми волокнами больше, чем безмиелиновыми. Тонкие волокна, бедные миелином, и безмиелиновые волокна проводят нервный импульс со скоростью 1-2 м/сек, тогда как толстые миелиновые - 5-120 м/сек.
6. Нервные окончания
Конечные разветвления нейронов образуют нервные окончания, которые контактируют с тканями (органами), воспринимающими нервные импульсы из центра, либо с тканями (органами), передающими воздействие внутренней и внешней среды и способствующими образованию нервных импульсов, направленных в центр. В связи с этим различают эффекторы и рецепторы.
Эффекторы, или двигательные и секреторные нервные окончания, проводят нервные импульсы от мозга к рабочему органу (мышца, железа) и регулируют его функцию, а рецепторы, или чувствительные нервные окончания, передают нервные импульсы, вызванные различными раздражителями извне (кожа, скелетная мускулатура, внутренние органы).
Эффекторы - двигательные и секреторные нервные окончания - имеют различное строение. На миоцитах неисчерченной мышечной ткани (сарколемма) они представлены в виде небольших концевых пуговок, из сплетения нейрофибрилл в виде петли, в центре которой находится скопление аксоплазмы. Субмикроскопически это окончание имеет структуру синапса (рис. 13) с типичными особенностями (см. ниже). Очень характерно, что одно нервное волокно, подходя к миоциту, делится на ветви, образующие подобие сплетения и оканчивающиеся концевыми пуговками на сарколемме. Этим увеличивается контакт нейрона с миоцитом. В месте контакта, как уже доказано, нервное окончание вырабатывает особые продукты обмена, называемые медиаторами (холин - и адренергические вещества), которые способствуют передаче импульса на рабочий орган (ткань, клетка), а также и сами могут быть причиной возбуждения и специфической работы.

Рис. 13. Схема субмикроскопического строения двигательного нервного окончания на миоците: 1 – постсиналтнческая мембрана миоцита; 2 – митохондрия; 3 – светлые пресинаптические пузырьки; 4 – пресинаптическая мембрана нейрита
Это проверяется введением их в кровь, лимфу, когда медиаторы проявляют свое действие на другие органы (ткани).
В связи с отмеченным, полагают, что увеличение степени разветвления одного нервного волокна для образования контакта с одной мышечной клеткой способствует выработке большого количества медиатора и лучшей передаче нервного импульса.
Методом перерезки при последующей дегенерации нервных волокон, о чем будет сказано позже, доказано, что неисчерченная мускулатура иннервируется вегетативной нервной системой (постганглионарные нервные волокна).
На сердечных миоцитах (мышечные клетки миокарда сердца) в основном наблюдаются такие же нервные окончания, как и на миоцитах неисчерченной мышечной ткани. Источником иннервации также является вегетативная нервная система, причем трудно отличить окончания постганглионарных симпатических и парасимпатических волокон.
Окончания на неисчерченной и сердечной исчерченной мышечной ткани являются экстрацеллюлярными (внеклеточными), а по утверждению Ж. Буке (1913), они проникают в саркоплазму миоцитов.
Скелетная мускулатура получает иннервацию от мозга, который посылает нервные волокна к мышечным волокнам.

Рис. 14. Схема субмикроскопического строения нейромышечного окончания: 1 – цитоплазма нейролеммоцита; 2 – ядро нейролеммоцита; 3 – нейролемма, переходящая в наружный слой сарколеммы; 4 – аксон; 5 – сарколемма; 6 – концевые веточки нервного волокна в продольном и поперечном сечениях; 7 – митохондрии в аскоплазме: 8 – первичное синаптическое пространство; 9 – саркосомы; 10 – вторичное синаптическое пространство; 11 – светлые пресинаптические пузырьки; 12 – пресинаптическая мембрана; 13 – постсинаптическая мембрана; 14 – ядро мышечного волокна; 15 – миофибрилла, состоящая из миофиламентов
Как показывает световой микроскоп, в месте контакта с мышечным волокном нервное волокно теряет миелиновую оболочку, эндоневрий сливается с сарколеммой, которая расщепляется на два листка и включает разветвление аксона в виде петелек и пластинок из нейрофибрилл и ядер нейролеммоцитов. Окончания на мышечных волокнах скелетной мускулатуры называются нейромышечными (концевые моторные - бляшками), которые заложены внутрь сарколеммы и изолированы своим основанием - подошвой - от саркоплазмы (М.Д. Лавдовский, 1884-1885; К.А. Лавров, 1941). Ноэль (1932) доказал, что митохондрии подошвы концевой моторной бляшки близки по структуре к наблюдаемой в нейролеммоцитах и резко разнятся от митохондрий исчерченного мышечного волокна. Кроме того, при некоторых патологических процессах отмечались (Л.И. Фалин, 1954; Е.К. Плечкова, 1955) реактивные изменения в ядрах нейролеммоцитов (размножение, вакуолизация) подошвы концевых моторных бляшек, а мышечные волокна оставались без видимых изменений.
На электронограммах также можно наблюдать контакт нервных и мышечных волокон в области концевых моторных бляшек. Субмикроскопически каждая из них (рис.14) состоит из нервного волокна, нейролемма которого сливается с сарколеммой, аксон образует нервный полюс, т.е. концевую веточку (пальцеообразные выросты) и вместе с пограничной мембраной мышечного волокна погружается в саркоплазму. Аксолемма окружает разветвления аксона и образует пресинаптическую мембрану с пальцеобразными выростами. Между аксоплазмой и примыкающей к ней пресинаптической мембраной располагается первичное синаптическое пространство, а между пальцеобразными складками той же мембраны - вторичное синаптическое пространство. Эти щелевидные пространства заполнены гомогенным веществом. Кнаружи от пресинаптической мембраны залегает цитолемма мышечного волокна, или постсинаптическая мембрана. В аксоплазме разветвлений аксона содержатся пресинаптические пузырьки и митохондрии, которые в отличие от наблюдаемых в исчерченном мышечном волокне имеют не поперечные, а продольные митохондриальные гребни.
Установлено, что в области концевой моторной бляшки (пресинаптической части) вырабатывается ацетилхолин, а в области ее подошвы (постсинаптической части) - ацетилхолинэстераза, способствующая гидролизу ацетилхолина (В.В. Португалов, 1955). Рядом экспериментов и наблюдений доказано, что концевые моторные бляшки обладают очень высокой реактивностью.
Эффекторы наблюдаются на клетках эпителия (нервно-эпителиальные окончания) и мезенхимы. На клетках реснитчатого эпителия заканчиваются постганглионарые нервные волокна симпатических звездчатых узлов (Лашков, 1948). Нервные волокна проникают в эпителий, оплетают каждую его клетку и заканчиваются на ней концевой пуговкой (пуговчатое утолщение) в месте расположения запирающей зоны (замыкающие пластинки). Подобные окончания образуются и на клетках железистого эпителия (К.А. Арнштейн, 1895; Н.И. Зазыбин, 1933). Это нервно-железистые окончания. Пуговчатыми окончаниями симпатических нервных волокон обладают также остеобласты, причем при их дифференцировке в остеоциты нервное окончание исчезает. На липоцитах и на элементах мезенхимы также установлены нервные окончания.
Все окончания на элементах соединительной ткани имеют трофическое значение.
Рецепторы (чувствительные нервные окончания) встречаются в двух основных формах: свободные и связанные с дополнительными образованиями (клетками).
Свободные нервные окончания образуются миелиновыми нервными волокнами, которые теряют все оболочки при вхождении в иннервируемую ткань. Они состоят из аксонов, переходящих в разветвления, заканчивающиеся концевыми колечками и пластинками, образованными нейрофибриллами и аксоплазмой. По предположению И.Н. Филимонова (1955), каждое разветвление одето цитоплазмой нейролеммоцитов. Отсюда понятие “свободные нервные окончания" является относительным. Эта форма нервных окончаний наблюдается в эпидермисе, во внешнем влагалище волоса, на волокнах неисчерченной и сердечной исчерченной мускулатуры и т.д.
В эпидермисе, например, свободные окончания образованы миелиновыми волокнами, которые подходят вплотную к базальной мембране, теряют миелин, разветвляются и заканчиваются концевыми пуговками и пластинками на клетках, причем эти разветвления не выходят за пределы блестящего слоя. В самых поверхностных слоях эпидермиса нервные нити приобретают варикозные расширения (вздутия) и фрагментируются, что свидетельствует о постоянном раздражении, воспринимаемом конечными разветвлениями описываемых волокон. Окончания в эпидермисе относятся к экстероцепторам (воспринимают болевые раздражения).
В наружном корневом влагалище волоса конечные разветвления аксона приобретают вид частокола, окружающего волос, т.е. параллельного его оси. Это окончания волосяных фолликулов.
Ко второй группе рецепторов относятся такие нервные образования, аксоны которых связаны со специальными осязательными (чувствительными) клетками, по-видимому, возникающими из нейролеммоцитов.
У всех млекопитающих в эпидермисе обнаружены осязательные мениски, которые впервые были найдены в пятачке свиньи (рис.15). Они состоят из светлых осязательных клеток овоидной или овоидно-эллиптической формы, которые лежат на нервных пластинках типа розеток. Осязательные мениски - типичные экстероцепторы (проводят тактильную чувствительность).

Рис. 15. Рецепторные нервные окончания в эпителии: А – кожи свиньи; Б – из эймерова органа крота; 1 – осязательные клетки; 2 – осязательные мениски; 3 – нервные волокна; 4 – сетчатая структура вокруг осязательной клетки
К описываемым рецепторам относятся и осязательные тельца продолговато эллиптической формы (рис.16). Они лежат в сосочковом слое кожи под эпидермисом перпендикулярно его поверхности и состоят из тонкой соединительной капсулы осязательного тельца. К ней подходит миелиновое нервное волокно, оно, как правило, под капсулой теряет миелиновую оболочку, разветвляется и заканчивается пластинками из нейрофибрилл, на которых лежат олигодендроглиоциты - чувствительные клетки. Осязательные тельца находят и в слизистых оболочках, например в надгортаннике.
Другой разновидностью второй группы рецепторных окончаний являются капсулированные нервные тельца. Они имеют сильно развитую капсулу и обладают разнообразным строением. До последнего времени сюда же относили и осязательные тельца.
Наиболее типичные капсулированные нервные тельца - это концевые нервные тельца, генитальные, пластинчатые тельца и др.

Рис. 16. Осязательные тельца: А – из сосочка кожи пальца человека; Б – из кожи пальца человека; 1 – эпидермис; 2 – основное волокно; 3 – добавочное волокно; 4 – расширение основного волокна; 5 – олигодендроглиоциты (осязательные клетки); 6 – веточка нервного волокна, выходящая из тельца; 7 – пластинка
Концевые нервные тельца (слизистые оболочки рта, языка, мочевыводящих путей) состоят из довольно толстой соединительнотканной капсулы, к которой подходит миелиновое нервное волокно, теряющее под капсулой миелин и в виде ленты или пластинки располагающееся в центре тельца. Нередко от пластинки отходят разветвления в виде шипиков. Пластинка окружается клетками типа нейролеммоцитов. В целом это так называемая внутренняя луковица.
Более сложно устроены генитальные тельца (слизистые оболочки половых органов, рта и дыхательного аппарата). Заметим, что такое название является неудачным, так как эти тельца не связаны со специфическими ощущениями от половых органов, как раньше предполагали. Это скорее аппараты тактильного чувства. Их внутренняя луковица состоит из разветвления в виде сетей и клубочков, образованных несколькими миелиновыми нервными волокнами, подходящими к тельцу.
 Широко распространены в организме пластинчатые тельца. Это почти макроскопические образования (сетчатый слой кожи, подкожная основа, брыжейка, паренхима многих желез - слюнных, поджелудочной, предстательной, надпочечника).
Широко распространены в организме пластинчатые тельца. Это почти макроскопические образования (сетчатый слой кожи, подкожная основа, брыжейка, паренхима многих желез - слюнных, поджелудочной, предстательной, надпочечника).

Рис. 17. Капсулированные нервные окончания: А – продольный разрез пластинчатого тельца из кожи человека; Б – поперечный срез пластинчатого тельца из кожи пальца человека; 1 – нервное волокно; 2 – внутренняя луковица; 3 – клетки пластинок капсулы
Тельца (рис. 17) состоят из очень толстой капсулы - наружной луковицы - результат наслоения нескольких десятков пластинок плотной соединительной ткани (или глии) с клетками и тканевой жидкостью между пластинками. Внутренняя луковица образована видоизмененными нейролеммоцитами и аксоном в виде длинной пластинки, иногда снабженной шипиками и выростами, а сам аксон ветвится. Кроме основного волокна, к тельцу подходит тонкое миелиновое или безмиелиновое добавочное волокно, аксон которого проникает во внутреннюю луковицу и разветвляется вокруг основного волокна. Раньше добавочные волокна относили к вегетативному отделу нервной системы, а в настоящее время (методом перерезки нервных волокон) установлена их цероброспинальная природа. Субмикроскопическое строение пластинчатых телец отличается от оптического некоторыми деталями (рис.18). По мнению большинства авторов, пластинчатые тельца реагируют на давление (проприоцепторы).

Рис. 18. Схема субмикроскопического строения пластинчатого тельца: 1 – слоистая капсула; 2 – спиральные коллагеновые волокна; 3– фиброцит; 4 – вторично чувствующие клетки с ресничками; 5 – дендрит рецепторного нейрона; 6 – внутренняя колба; 7 – синаптические контакты аксонов вторично чувствующих клеток с дендритами рецепторного нейрона
К капсулированным нервным окончаниям относят чувствительные концевые аппараты волокон скелетной мускулатуры - нервно-мышечные веретена. Строение их таково: одно или несколько очень тонких мышечных волокон заключено в специальную соединительно-тканную капсулу в форме овала или веретена. Толстые миелиновые нервные волокна прободают капсулу, теряют миелин, ветвятся в сопровождении специальных, по-видимому, глиальных клеток, подходят к мышечным волокнам и спирально их обвивают. Часть нервных нитей заканчивается спиральными пластинками на сарколемме мышечного волокна. Нервно-мышечные веретена передают в центр сигналы о состоянии тонуса мышечных волокон.
Исследованиями установлено наличие в кровеносной системе и в стенках ряда внутренних органов млекопитающих особых некапсулированных нервных окончаний - хеморецепторов, улавливающих изменения химического состава крови (общее сонное и грудное аортальное сплетения). Они состоят из разветвлений аксонов, сопровождаемых специальными клетками (глиальными).
Предполагают, что разные пороги раздражения общего сонного и грудного аортального сплетений объясняются разным количеством имеющихся в них специальных клеток.
В заключение следует отметить, что рецепторы обладают одной особенностью строения, изученной Б.И. Лаврентьевым (1941). Было установлено, что одно и то же чувствительное волокно на периферии ветвится и образует нервные окончания на различных тканевых образованиях разных органов, например в эпителии, в неисчерченных мышцах и на сосудах в желудочно-кишечном тракте, в сердечной исчерченной мускулатуре и на венечных сосудах. Такие окончания Б.И. Лаврентьев назвал “поливалентными" рецепторами. Нужно полагать, что к такому роду рецепторам относятся и те, которые способны к осуществлению аксон-рефлексов. Полагают, что аксон-рефлекс совершается через разветвления одного чувствительного нервного волокна, ветви которого связывают эпидермис с кровеносными сосудами. Раздражение эпидермиса вызывает расширение сосудов.
Положение Б.И. Лаврентьева о “поливалентных” рецепторах подтверждает правильность предположений авторов об аксон-рефлексе и определяет механизм его осуществления.
7. Синапсы
До последнего времени существовали две точки зрения о взаимосвязи между нейронами.
По мнению одних (Г. Гельд, 1895; К. Гольджи, 1880; С. Апати, 1897; А. Бете, 1903), использовавших материал беспозвоночных (особенно опыты Бете с крабом), нейрофибриллы - основной путь проведения нервных импульсов, а нейроцит - сгущение его цитоплазмы, имеющей трофическое значение. Было показано, что нейрофибриллы непрерывно переходят из одного нейрона в другой. Так возникла теория нейропиля (фибриллярной непрерывности, или нейрофибриллярной решетки). Применительно к высшим животным эта теория оказалась умозрительной, так как ее представителям не удалось убедить ученых в справедливости своего мнения из-за отсутствия демонстративных картин, типичных для беспозвоночных (крупные нервные клетки), что не типично для позвоночных, особенно высших (мелкие нервные клетки).

Рис.19.Схема строения синапса: 1 – пресинаптический полюс; 2 – митохондрии; 3 – светлый пресинаптический пузырек; 4 – пресинаптическая мембрана; 5 – синаптическая щель; 6 – Постсинаптическая мембрана; 7 – Постсинаптическая часть (на пре- и постсинаптической мембранах видны утолщения типа десмосом)
По мнению других (Р. Кахал, 1907; Б.И. Лаврентьев, 1939 и др.), между нейронами существует контакт - синапс (от греч. synapsis - соединение, связь). Термин дан английским физиологом Ч. Шеррингтоном. Впервые Р. Кахал установил синапсы в мозжечке: контактные связи разветвлений корзинчатых нейроцитов (вставочные нейроциты) с телами грушевидных нейроцитов (ганглиозный слой). Синапсы осуществляются передачей нервного импульса от нейрита чувствительного нейроцита к дендритам или телу корешкового (вставочного) нейроцита, а также от дендрита одного нейроцита к дендриту другого или от нейрита к нейриту двух нейроцитов. Кроме того, передача нервных импульсов может осуществляться от корешкового нейроцита к эпителиальным или мышечным клеткам.
Отсюда различают следующие синапсы: аксодендритические (контакт с дендритом последующего нейроцита), аксосоматические (контакт с телом последующего нейроцита), аксоаксональные (контакт между нейритами двух нейроцитов), дендродендритические (контакт между дендритами двух нейроцитов), аксоэпителиальные и аксомышечные (соответственно контакт корешковых нейроцитов с эпителио- и миоцитами).
Разные виды синапсов более или менее широко распространены в нервной системе. Так, аксосоматические синапсы имеют широкое распространение в нервной системе (корзинки нервных волокон на теле грушевидных нейроцитов, нейроны спинного, головного мозга, коры большого мозга, вегетативной нервной системы). Кроме того, известно, что нейрит одного нейрона, например чувствительного, может давать разветвления и оканчиваться на многих других нейронах (до 10000 синапсов). В свою очередь с одним и тем же нейроном, например корешковым нейроцитом, могут образовывать синапсы также много (1200-1800) нейронов.
Прижизненное существование синапсов у позвоночных впервые установлено в лаборатории Б.И. Лаврентьева на примере изучения внутрисердечных узлов амфибий (лягушка, жаба) с использованием метода, предложенного в 1939 г. Е.М. Граменицким (растянутая стенка предсердий на пробковом каркасе).
Прижизненными наблюдениями над синапсами и изменениями в них при тех или иных воздействиях, например ультракоротких волн, сопровождаемых возбуждением и как следствием этого желатинизацией, усилением гранулообразования (окраска янусгрюном, нейтральротом), доказано существование синапсов. Одновременно установлено совпадение наблюдаемого прижизненно и на фиксированных, а также окрашенных объектах. Субмикроскопически синапс состоит из пресинаптической, постсинаптической частей и синаптической щели. В пресинаптическую часть входит освобожденный от миелина аксон передающего нейрона, окруженный аксолеммой и содержащий в аксоплазме много митохондрий и до 3 млн. пресинаптических пузырьков диаметром 20-50 нм. В пузырьках находится медиатор, чаще всего ацетилхолин. Постсинаптическая часть представлена дендритом или телом воспринимающего нейрона, эпителиоцитом или миоцитом. Синаптическая щель расположена между пресинаптической и постсинаптической мембранами (рис. 19). Пресинаптическая мембрана - аксолемма передающего нейрона с пресинаптическим уплотнением. Элементы глии не внедряются в синаптическую щель. Постсинаптическая мембрана - цитолемма воспринимающего нейрона или другой клетки с постсинаптическим уплотнением. В ней находятся холинорецепторы (воспринимают действие ацетилхолина). Биохимическая активность в передаче нервного импульса принадлежит аксону передающего нейрона (пресинаптической части). Установлено, что перед передачей нервного импульса с нейрона на нейрон, при возбуждении передающего нейрона (пресинаптическая часть) пресинаптические пузырьки лопаются, медиатор изливается в синаптическую щель, возбуждает холинорецепторы постсинаптической мембраны и снижает ее электрический потенциал до критического. В результате этого в дендрит или тело воспринимающего нейрона (или другой клетки) усиленно проникают ионы натрия и в смежной зоне с постсинаптической мембраной возникает потенциал действия, т.е. нервный импульс. Передача нервных импульсов осуществляется всегда в одном направлении. Все это в значительной мере подтверждает справедливость взглядов представителей теории синаптических, контактных связей между нейронами. Доказано, что синаптические связи между нейронами (чувствительным, двигательным и др.) осуществляются при участии элементов нейроглии - синаптоглии (Куто, 1953). Это положение показывает, что в структуре и в функциональных проявлениях межнейронных синапсов и нервных окончаний имеются общие черты, так как и они, как правило, состоят из разветвлений аксонов, сопровождаемых нейроглией. Значение теории синапсов заключается в том, что она правильно объясняет полярность нейронов (направленность нервных импульсов от дендритов через тело к нейриту). Это положение подтверждается и данными о гистогенезе нервной ткани с установлением связи между нейронами и иннервируемыми ими тканями. Итак, нервная система едина, она состоит из нейронов и нейроглии, связанных между собой морфологически и функционально. Отсюда выдвинутая в свое время нейронная теория (В.М. Бехтерев, В. Гис, В. Вальдейер, Р. Кахал), которая трактовала, что нейрон - автономная единица нервной системы, не может быть признана вполне правильной. Рациональным в нейронной теории является то, что она подчеркивает функциональное значение нейрона в целом и в известной мере объясняет взаимоотношение центров и проводников в составе нервной системы. Накопившиеся в последующем данные позволяют внести дополнения в классическую нейронную теорию. Эти данные свидетельствуют о том, что нейрон и нейроглия взаимосвязаны, нейроны и глия очень реактивны к разнообразным внешним воздействиям и изменяются, но вместе с тем обладают значительными резервными возможностями для компенсации и восстановления нарушенных функций.
8. Понятия о рефлекторных дугах
Из материала, изложенного выше, следует, что нервная система организма состоит из большого количества отдельных нейронов. Нервные волокна пронизывают все ткани тела, образуя многочисленные рецепторные и эфферентные концевые аппараты.

Рис. 20. Простая рефлекторная дуга. 1 – чувствительная нервная клетка; 2 – рецептор в коже; 3 – дендрит чувствительной клетки; 4 – неврилемма; 5 – ядро леммоцита; 6 – миелиновыи слой; 7 – перехват нервного волокна; 8 – осевой цилиндр; 9 – насечка; 10 – аксон чувствительной клетки; 11 – двигательная клетка; 12 – дендриты двигательной клетки; 13 – аксон двигательной клетки; 14 – миелиновые волокна; 15 – эффектор; 16 – спинномозговой узел; 17 – дорсальная ветвь спинномозгового нерва; 18 – задний корешок; 19 – задний рог; 20 – передний рог; 21 – передний корешок; 22 – вентральная ветвь спинно-мозгового нерва.
Под влиянием процессов, протекающих в различных органах, и под воздействием внешних раздражений в чувствительных окончаниях возникают нервные импульсы, которые или непосредственно, или при участии ассоциативных нейронов, в том числе и нейронов центральной нервной системы, поступают в двигательные, эфферентные, нервные клетки и через их эфферентные окончания - ткани рабочих органов. Цепь нейронов, обеспечивающая проведение нервного импульса от рецептора чувствительного нейрона до двигательного окончания в рабочем органе, носит название рефлекторной дуги.
Если представить себе самую простую рефлекторную дугу, состоящую только из двух нейронов - чувствительного и двигательного, то путь нервного импульса будет выглядеть следующим образом: рецептор - дендрит - тело рецепторного нейрона - его аксон - синапс (контакт двух нейронов) - дендрит двигательной нервной клетки - ее тело - ее аксон - двигательное окончание (рис. 20). В подавляющем большинстве случаев между чувствительными и двигательными нейронами включены вставочные, или ассоциативные, нервные клетки.
У высших животных рефлекторные дуги состоят обычно из многих нейронов и, таким образом, имеют значительно более сложное строение, чем в приведенной схеме. Рефлекторные дуги могут быть “цереброспинального, соматического" типа и “автономного, вегетативного" типа.
Цереброспинальные рефлекторные дуги осуществляют главным образом произвольную регуляцию работы скелетной мускулатуры, тогда как автономные нервные дуги регулируют в основном непроизвольные сократительные движения гладкой мускулатуры внутренних органов.
9. Гистогенез нервной ткани
Гистогенез - единый комплекс координированных во времени и пространстве процессов пролиферации, дифференцировки, детерминации, интеграции и функциональной адаптации клеток (А.А. Клишов, 1970).
Под пролиферацией понимают рост и размножение тканевых клеток, при этом увеличивается не только их число, но и масса живого вещества.
В ходе развития тканевые клетки подвергаются дифференцировке, в результате чего они специализируются (накопление органелл специального назначения, например, миофибрилл и пр). Следствие этого - возникновение структурных и функциональных различий между клетками в составе ткани.
Под детерминацией большинство исследователей понимают определение пути развития клеток, состояние стойкого и необратимого закрепления результатов клеточной дифференцировки.
Соотношение между двумя этими процессами состоит в том, что в ходе дифференцировки клеток изменяется степень их детерминации. Различают лабильную, обратимую и стабильную, необратимую детерминацию. Это и определяет возникновение клеточных популяций.
В процессе гистогенеза по мере усиления дифференцировки тканевых клеток повышается степень их интеграции, так как дифференциация и интеграция составляют диалектическое единство процесса развития. В связи с интеграцией обращается внимание на межклеточные отношения, которые, по мнению некоторых, активизируют и определяют направление дифференцировки клеток. Большое значение в эволюции имеют и межклеточные отношения. Они обусловливают развитие различных частных функций.
Под функциональной адаптацией клеток развивающейся ткани понимают приспособление их к конкретным условиям функционирования, особенно в критические периоды эмбриогенеза, когда морфофункциональные свойства дифференцирующихся клеток изменяются. В результате наступают структурные и функциональные изменения и перестройка тканей, которые определяются степенью их онто - и филогенетической детерминации, различной дифференцировкой клеток, пролиферативной активностью, сложными межклеточными и межтканевыми взаимоотношениями в процессе развития и функционирования и пр.
Таким образом, описанные закономерности гистогенеза тесно связаны между собой и выражены процессами пролиферации, дифференциации, межклеточными и межтканевыми взаимоотношениями и функциональной адаптацией клеток. В одних случаях эти процессы могут быть ведущими, а в других - лишь частными проявлениями закономерностей гистогенеза.

Рис. 21. Формирование нервной трубки зародыша цыпленка. А–стадия нервной пластинки; Б–замыкание нервной трубки; В–обособление нервной трубки и ганглиозной пластинки от эктодермы; 1 – нервный желобок; 2 – нервные валики; 3 – кожная эктодерма; 4 –хорда; 5 – мезодерма; 6 – ганглиозная пластинка, 7 – нервная трубка; 8 – мезенхима
Нервная ткань развивается из дорсального утолщения эктодермы - нервной пластинки. В процессе развития зародыша нервная пластинка, прогибаясь, превращается сначала в нервный желобок, а затем замыкается в нервную трубку (рис.21) и обособляется от кожной эктодермы. Нервная трубка представляет собой эмбриональный зачаток всей нервной системы человека. Из нее в дальнейшем формируются головной и спинной мозг, а также периферические отделы нервной системы. При смыкании нервного желобка по бокам в области его приподнятых краев (нервных валиков) с каждой стороны выделяется группа клеток, которая по мере обособления нервной трубки от кожной эктодермы образует между нервными валиками и эктодермой сплошной слой - ганглиозную пластинку. Последняя служит исходным материалом для клеток чувствительных нервных узлов (спинальных и краниальных) и узлов вегетативной нервной системы, иннервирующей внутренние органы.

Рис. 22. Спинной мозг зародышей млекопитающих в разных стадиях развития: А – нервная пластинка; Б, В – участок нервной трубки в более поздней ста-дии развития; 1 – митоз клетки нервной пластинки; 2 – митоз в эпендим-ном слое; 3 – ядерный (плащевой) слой; 4 – наружный слой (краевая вуаль); 5 – внутренняя пограничная мембрана; 6 – наружная пограничная мембрана; 7 – мезенхима
Нервная трубка на ранней стадии своего развития состоит из одного слоя клеток цилиндрической формы, которые в дальнейшем интенсивно размножаются митозом и увеличиваются в количестве; в результате стенка нервной трубки утолщается (рис.22). В этой стадии развития в ней можно выделить три слоя: внутренний эпендимный слой, характеризующийся активным митотическим делением клеток; средний слой - мантийный (плащевой), клеточный состав которого пополняется как за счет митотического деления собственных клеток, так и путем перемещения их из внутреннего эпендимного слоя; наружный слой, называемый краевой вуалью. Последний слой образуется отростками клеток двух предыдущих слоев.
В дальнейшем клетки внутреннего слоя превращаются в цилиндрические эпендимные (глиальные) клетки, выстилающие центральный канал спинного мозга. Клеточные элементы мантийного слоя дифференцируются в двух направлениях. Из них возникают нейробласты, которые постепенно превращаются в зрелые нервные клетки, и спонгиобласты, дающие начало различным видам клеток нейроглии (астроцитам и олигодендроглиоцитам).

Рис. 23. Развитие нейробластов
По мере дифференцировки нейробласта изменяется субмикроскопическое строение его ядра и цитоплазмы. В ядре возникают участки различной электронной плотности в виде нежных зерен и нитей. В цитоплазме хорошо выявляются в большом количестве широкие цистерны и более узкие канальцы цитоплазматической сети, увеличивается количество рибосом. В нейробласте всегда хорошо развит пластинчатый комплекс. Первым доступным для наблюдения в световом микроскопе морфологическим признаком начавшейся дифференцировки нервных клеток следует считать возникновение в их цитоплазме тонких нейрофибрилл. Тело нейробласта постепенно приобретает грушевидную форму, а от его заостренного конца начинает развиваться отросток - аксон (рис.23). В дальнейшем нейробласты превращаются в зрелые нервные клетки - нейроны. Нейробласты и нейроны в период эмбрионального развития нервной системы делятся митозом.
Картину митоза и амитоза нейронов в центральной и периферической нервных системах можно иногда наблюдать и в постэмбриональный период. Размножаются нейроны и в условиях культивирования нервной ткани вне организма. В настоящее время возможность деления ряда нервных клеток можно считать установленной, хотя биологическое значение этого процесса требует дальнейшего изучения.
10. Регенерация нервной ткани
Способностью к размножению и к прогрессивному развитию в нервной системе взрослого организма могут обладать только элементы макроглии и микроглии. Нервные элементы, т.е. нейроны, целиком восстанавливаться не должны, так как для этого в нервной системе взрослого организма не остается необходимого камбиального источника. Но частичная регенерация в нервных элементах все же происходит. Она заключается в том, что в случае повреждения отростков того или иного нейрона последние могут при известных условиях восстанавливаться. Далее, если тело нейрона будет приведено в состояние сильного раздражения или выведено из состояния системных отношений, в котором оно находится в норме, характер уже существующих отростков может изменяться и могут появиться новые отростки. Таким образом, содержащая ядро часть нейрона, т.е. нервная клетка, сохраняет способность изменять свою форму, и восстанавливать утраченные части. Поэтому всякое повреждение в центральной нервной системе приводит, во-первых, к регенеративному раздражению глиальных и соединительнотканных элементов, образующих смешанный глиальносоединительнотканный рубец, и, во-вторых - к беспорядочному прорастанию этого рубца регенерирующими нервными отростками поврежденных нейронов. Ни восстановления утраченных нейронов, ни полного возобновления нарушенных связей при повреждении центральной нервной системы не происходит.
Наиболее изучены регенерация и дегенерация периферических нервов и составляющих их нервных волокон.
Нервные волокна представляют собой отростки нервных клеток и образуют с ними единую систему. Перерезка нервных волокон неизбежно вызывает изменения в теле нервных клеток, в центральном и периферическом отрезках волокна, а также реакцию со стороны нейроглии и окружающей соединительной ткани. Тело нейрона при этом увеличивается в объеме. Ядро несколько округляется и смещается на периферию клетки. Глыбки базофильного вещества постепенно исчезают. Тигролиз закономерно распространяется от ядра к периферии тела клетки. Этот процесс носит название центрального хроматолиза. Центральный отрезок волокна на некотором расстоянии от места травмы подвергается ретроградной, т.е. восходящей, дегенерации, распространяющейся от места травмы к телу клетки, после чего начинается его регенерация. Периферический отрезок подвергается вторичной (уоллеровской) дегенерации (рис. 191).
На месте перерезки возникает воспалительная реакция, в результате этого развивается нейроглиально-соединительнотканная рубцовая ткань - рубец, через который в дальнейшем будут прорастать центральные отрезки нервных волокон. Регенерация происходит тем быстрее, чем ближе расположены отрезки нерва и чем тоньше рубец.
Вторичная дегенерация периферического отрезка сводится к последовательным, связанным между собой изменениям осевого цилиндра и оболочки волокна. Осевой цилиндр в течение первых двух суток после перерезки несколько набухает, в результате чего по его ходу образуются значительные вздутия. В дальнейшем, на 3-5-е сутки, он распадается на фрагменты различной величины. Одновременно с этим изменяется и миелиновый слой оболочки волокна. Леммоциты резко активизируются. Уже в первые сутки после перерезки нервного волокна периферическая зона леммоцитов увеличивается в объеме. В отличие от нормальных волокон в условиях дегенерации в ней значительно усиливается цитоплазматическая сеть и увеличивается количество рибосом (рис. 192). Последние располагаются частично в виде полирибосом, частично они связаны с мембранами цитоплазматической сети. Одновременно перестраивается миелиновый слой оболочки волокна. Его мембраны теряют правильное, параллельное друг другу положение. Между группами мембран образуются значительные пространства. В дальнейшем мембраны фрагментируются и разрушаются. Миелиновый слой как обособленная зона леммоцита исчезает. В течение 3-4 суток леммоциты значительно увеличиваются в объеме. Субмикроскопическая структура их цитоплазмы, а именно плотная цитоплазматическая сеть, обилие рибосом, а позднее и митохондрий, свидетельствует о высокой функциональной активности леммоцитов. По мере распада мембран миелинового слоя в процессе дегенерации волокна в цитоплазме леммоцитов образуется значительное количество шарообразных слоистых структур различных размеров. Последние на микроскопических препаратах после обработки четырехокисью осмия выявляются в виде “капель миелина”. Цепочки леммоцитов на таких препаратах видны как плотные тяжи, в которых в особых вакуолях - “овоидах” - включена продукты распада миелина и осевых цилиндров. Леммоциты интенсивно (см. рис. 192) размножаются сначала амитозом, а затем кариокинезом. К концу второй недели миелин и частицы осевых цилиндров рассасываются. В резорбции продуктов распада принимают участие как глиальные клетки, так и макрофаги соединительной ткани.
Регенерация нервного волокна начинается с интенсивного размножения леммоцитов и образования ими лент (бюнгыеровские ленты), проникающих из периферического и центрального отрезков нерва в рубцовую ткань. Осевые цилиндры волокон центрального отрезка образуют на своих концах булавовидные расширения - колбы роста - и врастают в глиальный рубец, а позднее в бюнгнеровские ленты периферического отрезка нерва (рис. 193). Возможен рост осевых цилиндров и вне глиальных тяжей. Периферический нерв растет со скоростью 1-4 мм в сутки. Рост нервных волокон замедляется на периферии в области окончаний. Позднее образуется миелин, и волокно восстанавливает свой первоначальный характер.
Заключение
Мы рассмотрели гистогенез и регенерацию нервной ткани. И еще раз убедились, что нервная ткань самая специализированная и самая сложная ткань. Остается еще много вопросов, на которые невозможно ответить. Многие ученые заняты этой проблемой на разных уровнях и разных направлений. Нейробиологи, физиологи, медики, химики и биохимики (тем, кем я стану) бьются над нерешенными проблемами. На данном этапе бурно развиваются исследования головного мозга и способность к восстановлению нервных клеток. Получено множество данных, все это обобщается, но для развития необходим какой-то толчок, какое-то знание или методы получения, которых еще не изобретено. В будущем я хотел бы заниматься изучением проведения нервных импульсов и созданием на этой основе биокомпьютеров (вживленных в организм), способных выполнять заданные программы, а может и те, которые сами будут эволюционировать и развиваться?
Время покажет…
Литература
1. Альбертс Б., Брей Д. Молекулярная биология клетки М.: Мир, 1994. Т.3. С.287-374.
2. Антипчук Ю.П. Гистология с основами эмбриологии М.: Просвещение, 1988.240 с.
3. Березов Т.Т., Коровкин Б.Ф. Биологическая химия М.: Медицина, 1998. С.625-645.
4. Елисеев В.Г., Афанасьев Ю.И. Гистология М.: Медицина, 1972. С.224-249.
5. Заварзин А.А., Щелкунов С.И. Руководство по гистологии Л.: МедГИз, 1978. С.295-314.
6. Корочкин Л.И., Михайлов А.Т. Введение в нейрогенетику М.: Наука, 2000. С.7-98.
7. Мануилова Н.А. Гистология с основами эмбриологии М.: Просвещение, 1973.286 с.
8. Мецлер Д. Биохимия М.: Мир, 1980. Т.3. С.325-352
9. Николлс Дж.Г., Мартин А.Т. От нейрона к мозгу М.: УРСС, 2003. С.31-33, 143-163, 522-601.
10. Прохорова М.И. Нейрохимия Л.: Ленинградский университет, 1979. С.114-178
11. Рябов К.П. Гистология с основами эмбриологии Минск: Высшая школа, 1990.255 с.
12. Токин Б.П. Общая эмбриология М.: Высшая школа, 1987.474 с.
13. Уайт А., Хендлер Ф. Основы биохимии М.: Мир, 1981. Т.3. С.1425-1465.
14. Хухо Ф. Нейрохимия М.: Мир, 1990. С.91-107, 187-237.
15. Ченцов Ю.С. Введение в клеточную биологию М.: Наука, 1995.443 с.
16. Шеперд Г. Нейробиология М.: Мир, 1987. Т.1. С.35-71, 78-123.
17. Шульговский В.В. Основы нейрофизиологии М.: Аспект пресс, 2000. С.14-35.