| Скачать .docx |
Реферат: Белки, липиды и углеводы вирусов
Оглавление
1.Особенности строения вирусов. 6
2.Нуклеиновые кислоты вирусов. 11
3.Белки, липиды и углеводы вирусов. 22
Введение
Вирусы — это организмы, не способные существовать и размножаться самостоятельно. Они относятся к царству Vira.
В определении вируса подчеркивается особая природа их паразитизма, который можно назвать паразитизмом на генетическом уровне. Тот факт, что вирусы способны выживать и размножаться только внутри других клеток, объясняется не отсутствием собственной клеточной организации, а их потребностью в поступлении готовых источников питания. Если бактерии обладают способностью расти и размножаться на искусственных питательных средах, то вирусы, напротив, как настоящие клеточные паразиты, полностью зависят от обмена веществ в клетке-хозяине. Сейчас уже доказано, что отношение вирус—хозяин не ограничивается лишь питанием, а носит более сложный характер[1] .
Когда стали возможны современные методы исследования, с помощью электронного микроскопа удалось выявить детали структуры вирусов.
От бактерий вирусы отличаются простотой строения. Они состоят из нуклеиновой кислоты и белковой оболочки, которая называется «капсид».
Изучение вирусов началось в 1892 году, когда Д.И. Ивановский открыл вирус табачной мозаики. Вскоре, в 1898 году, была доказана вирусная природа ящура, а в 1917 году - были открыты бактериофаги. Вирусы сначала считали микроорганизмами, а бактериофаги - и вовсе разновидностью ферментов. В 60-х годах благодаря развивающейся молекулярной биологии, было введено понятие вириона - единицы вируса, и стало понятно, что вирусы не являются организмами. В 1962 году на 1-м Международном симпозиуме было сформулировано основное отличие вирусов от других живых организмов: генетический материал вирусов - или ДНК, или РНК, а организмы имеют оба типа нуклеиновых кислот. Другое главное отличие - отсутствие у вирусов собственных систем синтеза белка[2] .
Представители царства Вира не только не являются организмами, к ним неприменимы основные таксонометрические единицы биологии: особь, популяция, вид. Сначала казалось, что единица вируса - вирион - белковый кокон, с заключенной внутри вирусной ДНК или РНК. Но выяснилось, что один вирион содержит не все гены вируса, а лишь их фрагменты. Синтез новых вирусов начинается в зараженной клетке, когда в нее проникнут вирионы со всеми фрагментами генотипа - их может быть от 2 до 28. Потому нельзя сказать, что вирион - "вирусная особь". Но даже множество вирионов не образуют вирусную популяцию. Рядом с ними всегда будут так называемые дефектные вирусы и вирусы-саттелиты, - вирусы паразитирующие на другом вирусе. Дефектные вирусы размножаются в клетке только при наличии полноценного вируса-помощника (используя его гены, если дефект самого вируса - дефект гена полимеразы, или используя его белки - если у него дефект гена внутренних или оболочечных белков). При смешанных инфекциях генотип одного вируса находится в оболочке другого. Другой тип паразитирующих вирусов - саттелиты. Они используют полноценный неродственный вирус, и при репликации новый саттелит всегда имеет дефект в генотипе, не позволяющий ему самостоятельно размножаться[3] .
Есть разновидности вирусов, которые всегда находятся в клетке - плазмиды,оничасто выполняют полезные функции в клетках бактерий (например, синтезируют токсины, убивающие насекомых, или ферменты, разрушающие антибиотики), их репликация обеспечивается клеткой, а их гены не кодируют синтез белков. Прионы - возбудители спогиформных энцефалопатий (болезнь куру, болезнь Крейтцфельда-Якоби) - это результат выхода из-под контроля генов, кодирующих белки, в результате чего поражаются нервные клетки.
Целью данной работы является изучение биохимической структуры вирусов.
Для достижения поставленной цели в работе рассмотрим следующие задачи:
1. изучим общее строение вирусов;
2. на основе общего строения вирусов рассмотрим более детально нуклеиновый состав вирусов; строение белков, липидов и углеводов.
1. Особенности строения вирусов
Вирусы – это мельчайшие микробы (их размеры колеблются от 12 до 500 нанометров), не имеющие клеточного строения, белок-синтезирующей системы, содержащие только один тип нуклеиновой кислоты (ДНК или РНК).
Важнейшими отличительными особенностями вирусов являются следующие[4] :
1. Они содержат в своем составе только один из типов нуклеиновых кислот: либо рибонуклеиновую кислоту (РНК), либо дезоксирибонуклеиновую (ДНК), - а все клеточные организмы, в том числе и самые примитивные бактерии, содержат и ДНК, и РНК одновременно.
2. Не обладают собственным обменом веществ, имеют очень ограниченное число ферментов. Для размножения используют обмен веществ клетки-хозяина, ее ферменты и энергию.
3. Могут существовать только как внутриклеточные паразиты и не размножаются вне клеток тех организмов, в которых паразитируют.
Вирусы состоят из следующих основных компонентов:
1. Сердцевина - генетический материал (ДНК либо РНК), который несет информацию о нескольких типах белков, необходимых для образования нового вируса.
2. Белковая оболочка, которую называют капсидом (от латинского капса - ящик). Она часто построена из идентичных повторяющихся субъединиц - капсомеров. Капсомеры образуют структуры с высокой степенью симметрии.
3. Дополнительная липопротеидная оболочка. Она образована из плазматической мембраны клетки-хозяина и встречается только у сравнительно больших вирусов (грипп, герпес).
Капсид и дополнительная оболочка несут защитные функции, как бы оберегая нуклеиновую кислоту. Кроме того, они способствуют проникновению вируса в клетку. Полностью сформированный вирус называется вирионом.
Вирусы способны размножаться только в клетках других организмов. Вне клеток организмов они не проявляют никаких признаков жизни. Многие из них во внешней среде имеют форму кристаллов. Размеры вирусов колеблются в пределах от 20 до 300 нм в диаметре[5] .
Хорошо изучен вирус табачной мозаики, имеющий палочковидную форму и представляющий собой полый цилиндр. Стенка цилиндра образована молекулами белка, а в его полости расположена спираль РНК (рис. 1). Белковая оболочка защищает нуклеиновую кислоту от неблагоприятных условий внешней среды, а также препятствует проникновению ферментов клеток к РНК и ее расщеплению.
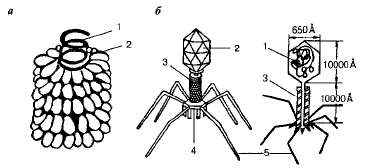
Рис. 1. Схема строения вируса (а) и бактериофага (б); 1— нуклеиновая кислота; 2 — белковая оболочка; 3 — полый стержень; 4 — базальная пластинка; 5 — отростки (нити).
Молекулы вирусной РНК могут самовоспроизводиться. Это означает, что вирусная РНК является источником генетической информации и одновременно иРНК. Поэтому в пораженной клетке в соответствии с программой нуклеиновой кислоты вируса на рибосомах клетки хозяина синтезируются специфические вирусные белки и осуществляется процесс самосборки этих белков с нуклеиновой кислотой в новые вирусные частицы. Клетка при этом истощается и погибает. При поражении некоторыми вирусами клетки не разрушаются, а начинают усиленно делиться, часто образуя у животных, в том числе и человека, злокачественные опухоли.
Поскольку основу всего живого составляют генетические структуры, то и вирусы классифицируют сейчас по характеристике их наследственного вещества - нуклеиновых кислот. Все вирусы подразделяют на две большие группы: ДНК-содержащие вирусы (дезоксивирусы) и РНК-содержащие вирусы (рибовирусы). Затем каждую из этих групп подразделяют на вирусы с двухнитчатой и однонитчатой нуклеиновыми кислотами. Следующий критерий - тип симметрии вирионов (зависит от способа укладки капсомеров), наличие или отсутствие внешних оболочек и т.п.
Ниже в таблице 1 представлена современная классификация вирусов и в качестве примера приведены наиболее известные вирусы.
Таблица 1.
Классификация вирусов . |
|||
| ДЕЗОКСИВИРУСЫ | РИБОВИРУСЫ | ||
| 1. ДНК двухнитчатая | 2. ДНК однонитчатая | 1. РНК двухнитчатая | 2. РНК однонитчатая |
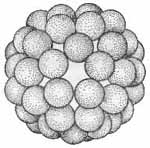
1.1. Кубический тип симметрии : 1.1.1. Без внешних оболочек: аденовирусы (см рис 2в ) 1.1.2. С внешними оболочками: герпес-вирусы(см рис 2б ) 1.2. Смешанный тип симметрии : Т-четные бактериофаги (см.рис 1) 1.3. Без определенного типа симметрии : оспенные вирусы |
2.1. Кубический тип симметрии : 2.1.1. Без внешних оболочек: крысиный вирус Килхама, аденосателлиты |
1.1. Кубический тип симметрии : 1.1.1. Без внешних оболочек: реовирусы, вирусы раневых опухолей растений |
2.1. Кубический тип симметрии : 2.1.1. Без внешних оболочек: вирус полиомиелита (см.рис 2г ), энтеровирусы, риновирусы 2.2. Спиральный тип симметрии : 2.2.1. Без внешних оболочек: вирус табачной мозаики 2.2.2. С внешними оболочками: вирусы гриппа(см рис 2а ), бешенства, онкогенные РНК-содержащие вирусы |
Приведенная таблица имеет некоторое сходство с таблицей Менделеева. В ней тоже есть незаполненные места. Так, например, до сих пор неизвестны дезоксивирусы со свойствами 2.2 (однонитчатая ДНК, спиральный тип симметрии) или рибовирусы со свойствами 1.2 (РНК двухнитчатая, смешанный тип симметрии). Может быть, что таких вирусов и нет в природе, а может, их еще не открыли. Совсем недавно рибовирусы со свойствами 1.1.1 не были известны, но затем оказалось, что к ним относятся реовирусы и сходные с ними вирусы раневых опухолей растений. То же самое относится и к дезоксивирусам со свойствами 2.1.1.
Ближайшие годы покажут, реализовала ли природа все возможные схемы строения вирусов, или некоторые из них оказались нежизненными и потому нереализованными.
|
 |
| а | б |
 |
 |
в |
г |
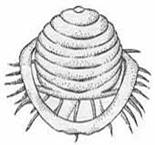
Рис. 2. Схематичное изображение расположения капсомеров в капсиде вирусов. Спиральный тип симметрии имеет вирус гриппа - а . Кубический тип симметрии у вирусов: герпеса - б , аденовируса - в , полиомиелита - г .
Из таблицы классификации вирусов видно, что разнообразие в царстве вирусов значительно более выражено, нежели в растительном и животном мире, если за основу взять характеристику генетических структур. В самом деле, все животные и растения - от амебы до человека и от бактерии до цветкового растения - имеют генетический материал в виде двухнитчатой ДНК. У вирусов же генетическим материалом могут быть однонитчатые и двухнитчатые формы обеих нуклеиновых кислот.
2. Нуклеиновые кислоты вирусов
В отличие от клеток вирусы содержат лишь один вид нуклеиновой кислоты – либо РНК, либо ДНК. И та, и другая может быть хранителем наследственной информации, выполняя таким образом функции генома.
Вирусные нуклеиновые кислоты характеризуются поразительным разнообразием форм. Вирусный геном может быть представлен как однонитчатым, так и двунитчатыми молекулами РНК и ДНК. ДНК может быть как линейной, так и кольцевой молекулой, РНК – как непрерывной, так и фрагментированной и кольцевой молекулой (рис.3).
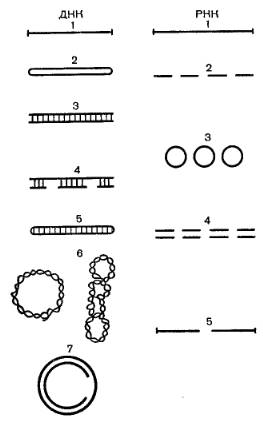
Рис.3. типы молекул вирусных ДНК и РНК.
ДНК: 1 – парвовирусов; 2 – фага XI74; 3 – аденовирусов, вирусов герпеса; 4 – фага Т5; 5 – вирусов оспы; 6 – паповавирусов; 7 – вируса гепатита В.
РНК: 1 – пикорнавирусов, тогавирусов, парамиксовирусов, рабдовирусов; 2 – ортомиксовирусов, аренавирусов; 3 – буньявирусов; 4 – реовирусов; 5 – ретровирусов.
Геном вирусов включает[6] :
- Структурные гены, которые кодируют белки. Занимают примерно 95 % вирусной хромосомы. Белки вирусов можно разделить на несколько групп: структурные, ферменты, регуляторы.
- Регуляторные последовательности, которые не кодируют белки: промоторы, операторы и терминаторы.
- Прочие некодирующие участки (сайты), в том числе:
- участок attP , обеспечивающий интеграцию вирусной хромосомы в хромосому клетки–хозяина;
- участки cos – липкие концевые участки линейных вирусных хромосом, обеспечивающие замыкание линейной хромосомы в кольцевую форму.
Гены, кодирующие рРНК и тРНК, в геноме вирусов обычно отсутствуют. Однако в геноме крупного фага Т4 имеются гены, кодирующие несколько тРНК.
Геном вирусов отличается высокой плотности упаковки информации. Например, у фага φ Х174 в пределах одного гена может располагаться еще один ген (на рисунке кольцевая ДНК представлена в линейной форме). В частности, ген В находится в пределах гена А , а ген Е – в пределах гена D :
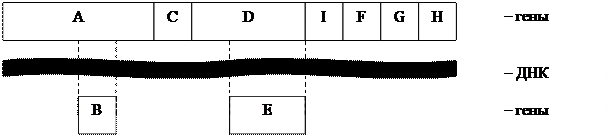 |
У мелкого РНК–содержащего фага f2 ген регуляторного белка, блокирующего лизис (созревание вирионов и разрушение клетки), перекрывается с двумя другими генами, удаленными друг от друга:
 |
Экспрессия (транскрипция и трансляция) вирусных генов происходит в том случае, если геном вируса представлен двунитевой ДНК (у РНК–содержащих вирусов необходим перевод информации в ДНК). Из-за полярности ДНК транскрипция идет только в одном направлении, то есть ген имеет начало и конец. Тогда «правые» гены не будут транскрибироваться РНК–полимеразой, движущейся влево, и наоборот. При этом один и тот же ген может транскрибироваться с разных промоторов; в этом случае экспрессия генов контролируется разными механизмами[7] .
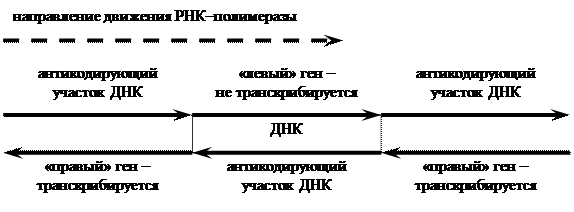 |
2.1. Вирусные ДНК
Молекулярная масса ирусных ДНК варьирует в широких пределах от 1×106 до 250×106 . Самые большие вирусные геномы содержат несколько сотен генов, а самые маленькие содержат информацию, достаточную для синтеза лишь несколько белков.
В геномах, представленных двунитчатыми ДНК, информация обычно закодирована на обеих нитях ДНК. Это свидетельствует о максимальной экономии генетического материала у вирусов, что является неотъемлемым свойством их как генетической информации не может быть проведена по молекулярной массе молекул[8] .
Хотя в основном структура ДНК уникальна, т.е. большинство нуклеотидных последовательностей встречаются лишь по одному разу, однако на концах молекул имеются повторы, когда в концевом фрагменте линейной ДНК повторяется ее начальный участок. Повторы могут быть прямыми и инвертированными[9] .
Способность к приобретению кольцевой формы, которая потенциально заложена в концевых прямых и инвертированных повторах, имеет большое значение для вирусов. Кольцевая форма обеспечивает устойчивость ДНК к экзонуклеазам. Стадия образования кольцевой формы обязательна для процесса интеграции ДНК с клеточным геномом. Наконец, кольцевые формы представляют собой удобный и эффективный способ регуляции транскрипции и репликации ДНК.
В составе вирионов, содержащих однонитчатую ДНК, обычно содержатся молекулы ДНК одной полярности. Исключение составляют адноассоциированные вирусы, вирионы которых содержат ДНК либо одной полярности (условно называемой «плюс»), либо ДНК с противоположным знаком (условно «минус»). Поэтому тотальный препарат вируса состоит из двух типов частиц, содержащих по одной молекуле «плюс»- или «минус»-ДНК.
Инфекционный процесс при заражении этими вирусами возникает лишь при проникновении в клетку частиц обоих типов.
Первый этап репликации вирусов связан с проникновением вирусной нуклеиновой кислоты в клетку организма-хозяина. Этому процессу могут способствовать специальные ферменты, входящие в состав капсида или внешней оболочки вириона, причем оболочка остается снаружи клетки или вирион теряет ее сразу после проникновения внутрь клетки. Вирус находит подходящую для его размножения клетку, контактируя отдельными участками своего капсида (или внешней оболочки) со специфическими рецепторами на поверхности клетки по типу «ключ – замок». Если специфические («узнающие») рецепторы на поверхности клетки отсутствуют, то клетка не чувствительна к вирусной инфекции: вирус в нее не проникает[10] .
Для того чтобы реализовать свою генетическую информацию, проникшая в клетку вирусная ДНК транскрибируется специальными ферментами в мРНК. Образовавшаяся мРНК перемещается к клеточным «фабрикам» синтеза белка – рибосомам, где она заменяет клеточные «послания» собственными «инструкциями» и транслируется (прочитывается), в результате чего синтезируются вирусные белки. Сама же вирусная ДНК многократно удваивается (дуплицируется) при участии другого набора ферментов, как вирусных, так и принадлежащих клетке.
Синтезированный белок, который используется для строительства капсида, и размноженная во многих копиях вирусная ДНК объединяются и формируют новые, «дочерние» вирионы. Сформированное вирусное потомство покидает использованную клетку и заражает новые: цикл репродукции вируса повторяется. Некоторые вирусы во время отпочковывания от поверхности клетки захватывают часть клеточной мембраны, в которую «заблаговременно» встроились вирусные белки, и таким образом приобретают оболочку. Что касается клетки-хозяина, то она в итоге оказывается поврежденной или даже полностью разрушенной[11] .
У некоторых ДНК-содержащих вирусов сам цикл репродукции в клетке не связан с немедленной репликацией вирусной ДНК; вместо этого вирусная ДНК встраивается (интегрируется) в ДНК клетки-хозяина. На этой стадии вирус как единое структурное образование исчезает: его геном становится частью генетического аппарата клетки и даже реплицируется в составе клеточной ДНК во время деления клетки. Однако впоследствии, иногда через много лет, вирус может появиться вновь – запускается механизм синтеза вирусных белков, которые, объединяясь с вирусной ДНК, формируют новые вирионы[12] .
2.2. Вирусные РНК
Из нескольких сотен известных в настоящее время вирусов человека и животных РНК-геном содержит около 80% вирусов. Способность РНК хранить наследственную информацию является уникальной особенностью вируса.
У просто организованных и некоторых сложно организованных вирусов вирусная РНК в отсутствие белка может вызвать инфекционный процесс. Впервые инфекционная активность РНК вируса табачной мозаики была продемонстрирована Х.Френкель-Конратом и соавт. в 1957г. и А. Гирером и Г.Шраммом в 1958г. Впоследствии положение об инфекционной активности РНК было перенесено на все РНК-содержащие вирусы, однако долголетние усилия доказать это для таких вирусов, как вирусы гриппа, парамиксовирусы, рабдовирусы (так называемые минус-нитевые вирусы), оказались бесплодными: у этих вирусов инфекционной структурой являются не РНК, а комплекс РНК с внутренними белками. Таким образом, геномная РНК может обладать инфекционной активностью в зависимости от своей структуры[13] .
Структура вирусных РНК чрезвычайно разнообразна. У вирусов обнаружены однонитчатые и двунитчатые, линейные, фрагментированные и кольцевые РНК. РНК-геном в основном является гаплоидным, но геном ретровирусов – диплоидный, т.е. состоит из двух идентичных молекул РНК.
Однонитчатые РНК. Молекулы однонитчатых вирусных РНК существуют в форме одиночной полинуклеотидной цепи со спирализированными ДНК-подобными участками. При этом некомплементарные нуклеотиды, разделяющие комплементарные участки, могут выводиться из состава сперализованных участков в форме различных «петель» и «выступов» (рис.4). Суммарный процент спирализации вирусных РНК не обнаруживает каких-либо особенностей по сравнению с таковыми у клеточных РНК.
Вирусы, содержащие однонитчатые РНК, делятся на две группы. У вирусов первой группы вирусный геном обладает функциями информационной РНК, т.е. может непосрдственной переносить закодированную в нем информацию на рибосомы. По предложению Д.Балтимора (1971), РНК со свойствами информационной условно обозначена знаком «плюс» и в связи с этим вирусы, содержащие такие РНК (пикорнавирусы, тогавирусы, коронавирусы, ретровирусы), обозначены как «плюс-нитевые» вирусы, или вирусы с позитивным геномом[14] .
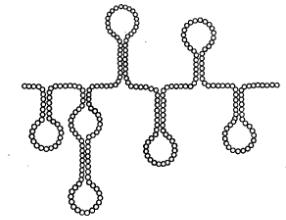
Рис. 4. Вторичная структура вирусных РНК (схема).
Вторая группы РНК-содержащих вирусов содержит геном в виде однонитчатой РНК, которая сама не обладает функциями иРНК. В этом случае функцию иРНК выполняет РНК, комплементарная геному. Синтез этой РНК (транскрипция) осуществляется в зараженной клетке на матрице геномной РНК с помощью вирусспецифического фермента – транскриптазы. В составе «минус-нитевых» вирусов обязательно присутствие собственного фермента, осуществляющего транскрипцию геномной РНК и синтез иРНК, т.к. аналога такого фермента в клетках нет. Геном этих вирусов условно обозначают как «минус»-РНК, а вирусы этой группы как «минус-нитевые» вирусы, или вирусы с негаивным геномом. К этим вирусам относятся ортомиксовирусы, парамиксовирусы, буньявирусы, рабдовирусы. РНК этих вирусов не способна вызвать инфекционный процесс.
В соответствии с разными свойствами вирусных РНК между двумя группами вирусов есть и структурные различия. Поскольку РНК «плюс-нитевых» вирусов выполняет функцию иРНК, она имеет специфические структурные особенности, характерные для 5¢- и 3¢-концов этих РНК.
5¢-Конец клеточных и вирусных РНК обычно имеет структуру так называемой шапочки (по-английски «сар»):
![]()
где m 7 G представляет собой 7-метилгуанин, присоединенный через пирофосфатную связь к гуаниловому нуклеотиду, сахарный остаток которого также метилирован по второму углеродному атому. На 3¢-конце информационных РНК имеются поли(А), количество которых достигает 200 и выше. Эти модификации концов иРНК, осуществляемые после синтеза полинуклеотидной цепи, имеют существенное значение для функции иРНК: «шапочка» нужна для специфического узнавания РНК рибосомами, функции поли(И) менее точно определены и, по-видимому, заключается в придании стабильности молекулам иРНК.
Такими же модифицированными концами обладают геномные РНК «плюс-нитевых» вирусов. Исключение составляет 5¢-конец геномной РНК вируса полиомиелита, которая не содержит «шапочку», и вместо нее имеет на 5¢-конце ковалентно присоединенный к остатку урацила низкомолекулярный терминальный белок. Геномные РНК «минус-нитевых» вирусов не имеют ни «шапочки», ни поли(А); модифицированные концы характерны для иРНК этих вирусов, синтезирующихся в клетке на матрице вирионной РНК и комплементарных ей. Геномная РНК ретровирусов, хотя и являетс «плюс-нитевой», однако не содержит «шапочку»; эту структуру содержит гомологичная РНК, синтезируемая на матрице интегрированной провирусной ДНК.
Существуют вирусы, содержащие как «плсю-нитевые», так и «минус-нитевые» РНК гены (амбисенс-вирусы). В ним относятся аренавирусы.
В основном однонитчатые РНК являются линейными молекулами, однако РНК-фрагменты буньявирусов обнаружены в виде кольцевой формы. Кольцевая форма возникает за счет образования водородных связей между концами молекул.
Двунитчатые РНК . Этот необычный для клетки тип нуклеиновой кислоты, впервые обнаруженный у реовирусов, широко распространен среди вирусов животных, растений и бактерий. Вирусы, содежащие подобный геном, называют диплорнавирусы.
Общей особенностью диплорнавирусов является фрагментированное состояние генома. Так, геном реовирусов состоит из 10 фрагментов, ротавирусов – из 11 фрагментов[15] .
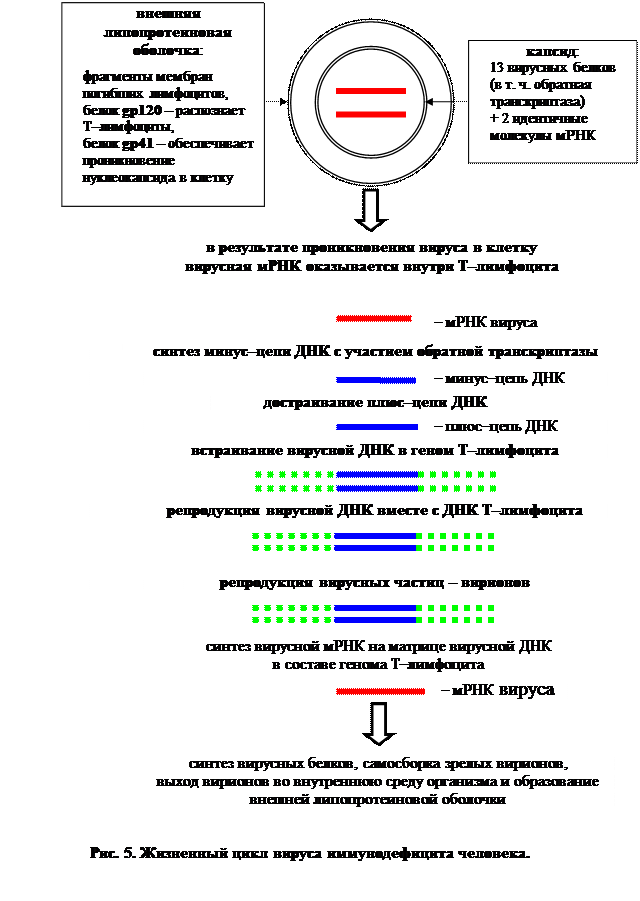
Рассмотрим процесс заражения на примере ВИЧ-инфицирования.
ВИЧ относится к ретровирусам. Его геном представлен линейной одноцепочечной молекулой РНК (это плюс–цепь, или мРНК). ВИЧ – это сложный вирус. В состав вириона входит нуклеокапсид (собственно вирион) и внешняя липопротеиновая оболочка. В состав нуклеокапсида входит РНК (две молекулы) и 13 вирусных белков, в том числе, и обратная транскриптаза (ревертаза). В состав липопротеиновой оболочки вириона входят фрагменты мембран погибших лимфоцитов, содержащие комплекс из двух особых белков: гликопротеидов gp41 и gp120. Белок gp120 распознает Т–лимфоциты, а белок gp41 «пробивает» мембрану Т–лимфоцита и обеспечивает проникновение нуклеокапсида в клетку[16] .
В инфицированной клетке на матрице вирусной РНК с помощью вирусной обратной транскриптазы синтезируется минус–цепь ДНК, а на ней – плюс–цепь ДНК. Вирусная РНК при этом разрушается. Образовавшаяся двухцепочечная ДНК встраивается в определенный участок одной из хромосом клетки хозяина. Интегрированная ДНК-копия вирусного генома представляет собой провирус . В таком состоянии ретровирус может долгое время сосуществовать с инфицированным организмом – вирусоносителем. Однако, получив определенные молекулярные сигналы, с провирусной ДНК инициируется транскрипция вирусной мРНК, а с нее – синтез вирусных белков. Затем происходит самосборка множества вирусных частиц и их выход из клетки. Клетка хозяина (лимфоцит) погибает, а часть ее мембран образует липопротеиновую оболочку вируса. Гибель Т-хелперов и приводит к развитию СПИДа.
3. Белки, липиды и углеводы вирусов
3.1. Белки вирусов
Количество структурных белков в составе вирусной частицы варьирует в широких пределах в зависимости от сложности организации вириона. Наиболее просто организованный вирус табачной мозаики содержит всего один небольшой белок с молекулярной массой 17—18- 103 , некоторые фаги содержат 2—3 белка, просто организованные вирусы животных — 3—4 белка. Сложно устроенные вирусы, такие как вирусы оспы, содержат более 30 структурных белков[17] .
Структурные белки делятся на 2 группы:
1) капсидные белки, образующие капсид, т. е. футляр для нуклеиновой кислоты вируса (от лат. сарза — вместилище), и входящие в состав капсида геномные белки, и ферменты;
2) суперкапсидные белки, входящие в состав суперкапсида, т. е. наружной вирусной оболочки.
Поскольку суперкапсид называют также «пеплос», эти белки называют пепломерами.
Просто организованные вирусы, представляющие собой нуклеокапсид, содержат только капсидные белки. Сложно организованные вирусы содержат капсидные и суперкапсидные белки.
Капсидные белки . Первоначальное представление о том, что капсидные белки являются всего лишь инертной оболочкой для вирусной нуклеиновой кислоты, сложилось на основании изучения наиболее просто организованного вируса табачной мозаики, частица которого состоит из одной молекулы РНК и одного типа белка, образующего чехол для РНК. Однако такое представление неправильно. Хотя основной функцией капсидных белков является функция защиты вирусного генома от неблагоприятных воздействий внешней среды, у многих вирусов в составе капсида есть белки и с другими функциями. Поэтому термин «капсид» далеко выходит за пределы представления о нем как о футляре или чехле для вирусной нуклеиновой кислоты.
В составе капсида некоторых вирусов (пикорнавирусы, паповавирусы, аденовирусы) содержатся белки, ковалентно связанные с вирусным геномом (геномные белки). Эти белки являются терминальными, т. е. соединенными с концом вирусной нуклеиновой кислоты. Функции их неразрывно связаны с функциями генома и их регуляцией.
У ряда сложно организованных вирусов в составе капсида имеются ферменты, осуществляющие транскрипцию и репликацию вирусного генома — РНК и ДНК (РНК- и ДНК-полимеразы), а также ферменты, модифицирующие концы иРНК. Если ферменты и геномные белки представлены единичными молекулами, то капсидные белки представлены множественными молекулами. Эти белки и формируют капсидную оболочку, в которую у сложно организованных вирусов вставлены молекулы белков с другими функциями.
Основным принципом строения капсидной оболочки вирусов является принцип субьединичности, т. е. построение капсидной оболочки из субъединиц-капсомеров, образованных идентичными полипептидными цепями. Правильно построенные белковые субъединицы — капсомеры возникают благодаря способности вирусных капсидных белков к самосборке. Самосборка объясняется тем, что упорядоченная структура — капсид имеет наименьшую свободную энергию по сравнению с неупорядоченными белковыми молекулами. Сборка капсидной оболочки из субъединиц запрограммирована в первичной структуре белка и происходит самопроизвольно или при взаимодействии с нуклеиновой кислотой.
Суперкапсидные белки . Гликопротеиды. Суперкапсидные белки, или пепломеры, располагаются в липопротеидной оболочке (суперкапсиде или пеплосе) сложно устроенных вирусов. Они либо пронизывают насквозь липидный бислой как, например, гликопротеиды альфа-вирусов (вируса леса Семлики), либо не доходят до внутренней поверхности. Эти белки являются типичными внутримембранными белками и имеют много общего с клеточными мембранными белками. Как и последние, суперкапсидные белки обычно гликозилированы. Углеводные цепочки прикреплены к молекуле полипептида в определенных участках. Гликозилирование осуществляют клеточные ферменты, поэтому один и тот же вирус, продуцируемый разными видами клеток, может иметь разные углеводные остатки: может варьировать как состав углеводов, так и длина углеводной цепочки и место прикрепления ее к полипептидному остову[18] .
У большинства вирусов гликопротеиды формируют «шипы» на поверхности вирусной частицы, длина которых достигает 7—10 им. Шипы представляют собой морфологические субъединицы, построенные из нескольких молекул одного и того же белка. Вирусы гриппа имеют два типа шипов, построенных соответственно из гемагглютинина и нейраминидазы. Парамиксовирусы также имеют два типа шипов, построенных соответственно из двух гликопротеидов (НИ и Р), рабдовирусы имеют только один гликопротеид и, соответственно, один тип шипов, а альфа-вирусы имеют два или три гликопротеида, формирующих один тип шипов.
Гликопротеиды являются амфипатическими молекулами: они состоят из наружной, гидрофильной части, которая содержит на конце аминогруппу ДО -конец), и погруженной в липидный бислой, гидрофобной части, которая содержит на погруженном конце гидроксильную группу (С-конец). С-концом полипетид «заякоривается» в липид ном бислое. Есть, однако, и исключения из этого общего положения: нейраминидаза вируса гриппа взаимодействует с липидным бислоем не С-, а N-концом.
Основной функцией гликопротеидов является взаимодействие со специфическими рецепторами клеточной поверхности. Благодаря этим белкам осуществляется распознавание специфических клеточных рецепторов и прикрепление к ним вирусной частицы, т. е. адсорбция вируса на клетке. Поэтому гликопротеиды, выполняющие эту функцию, называют вирусными прикрепительными белками.
Другой функцией гликопротеидов является участие в слиянии вирусной и клеточной мембран, т. е. в событии, ведущем к проникновению вирусных частиц в клетку. Вирусные белки слияния ответственны за такие процессы, как гемолиз и слияние плазматических мембран соседних клеток, приводящие к образованию гигантских клеток, синци-тиев и симпластов.
«Адресная функция» вирусных белков. Вирусы вызывают инфекционный процесс у относительно небольшого круга хозяев. Вирус должен «узнать» чувствительную клетку, которая сможет обеспечить продукцию полноценного вирусного потомства. Если бы вирус проникал в любую клетку, которая встретилась на его пути, это привело бы к исчезновению вирусов в результате деструкции "«родительской» вирусной частицы и отсутствия вирусного потомства. В процессе эволюции у вирусов вырабатывалась так называемая адресная функция, т. е. поиск чувствительного хозяина среди бесконечного числа нечувствительных клеток. Эта функция реализуется путем наличия специальных белков на поверхности вирусной частицы, которые узнают специфический рецептор на поверхности чувствительной клетки.
Неструктурные белки . Неструктурные белки изучены гораздо хуже, чем структурные, поскольку их выделяют не из очищенных препаратов вирусов, а из зараженных клеток, и возникают трудности в их идентификации и очистке от клеточных белков.
К неструктурным белкам относятся:
1) предшественники вирусных белков, которые отличаются от других неструктурных белков нестабильностью в зараженной клетке в результате быстрого нарезания на структурные белки;
2) ферменты синтеза РНК и ДНК (РНК- и ДНК-26 полимеразы), обеспечивающие транскрипцию и репликацию вирусного генома;
3) белки-регуляторы;
4) ферменты, модифицирующие вирусные белки, например протеиназы и протеинкиназы.
Однако многие неструктурные белки при ряде вирусных инфекций еще не идентифицированы и функции их не определены. Типы структурных и неструктурных белков просто и сложно устроенных вирусов и их функции показаны на схеме (рис.6).
![]()
| Капсидные | |
| Геномные | |
| Неструктурные | |
| Функции не идентифицированы |
| Капсидные | |||
| Геномные | |||
| Полимеразы | Неструктурные | ||
| В составе суперкапсида | Белки слияния | Функции не идентифицированы | |
| Ферменты, модифицир- ующие белки | |||
| Прикрепительные |
Рис. 6. Типы структурных и неструктурных белков просто и сложно устроенных вирусов.
3.2. Липиды вирусов
Липиды обнаружены у сложно организованных вирусов и в основном находятся в составе липопротеиднои оболочки (суперкапсида), формируя ее липидной бислой, в который вставлены суперкапсидные белки.
Все сложно организованные РНК-содержащие вирусы имеют в своем составе значительное количество липидов (от 15 до 35% от сухого веса). Из ДНК-содержащих вирусов липиды содержат вирусы оспы, герпеса и гепатита В. Примерно 50—60% липидов в составе вирусов представлено фосфолипидами, 20—30% составляет холестерин.
Липидный компонент стабилизирует структуру вирусной частицы. Экстракция липидов органическими растворителями, обработка вирусной частицы детергентами или липазами приводит к деградации вирусной частицы и потере инфекционной активности[19] .
Вирусы, содержащие липопротеидную мембрану, формируются путем почкования на плазматической мембране клеток (или на мембранах эндоплазматической сети с выходом во внутриклеточные вакуоли). Поэтому липопротеидная оболочка этих вирусов представляет собой мембрану клетки-хозяина, модифицированную за счет наличия на ее наружной поверхности вирусных супер-капсидных белков. Из этого следует, что состав липидов почкующихся вирусов близок к составу липидов клетки-хозяина. К почкующимся вирусам относятся крупные
РНК-содержащие вирусы: ортомиксовирусы, парамиксо
вирусы, рабдовирусы, тогавирусы, ретровирусы, бунья
вирусы, аренавирусы, коронавирусы.
В связи с клеточным происхождением липидов общий состав липидной фракции и содержание ее отдельных компонентов у одного и того же вируса могут существенно различаться в зависимости от клетки-хозяина, где происходила репродукция вируса. Наоборот, если разные почкующиеся вирусы репродуцировались в одних и тех же клетках, их липиды оказываются более или менее сходными[20] .
У вирусов оспы и гепатита В липиды имеют иное происхождение, так как эти вирусы не почкуются через плазматическую мембрану. У вирусов оспы липиды не образуют дифференцированной оболочки. Обработка вируса осповакцины эфиром не приводит к потере инфекционной активности или каким-либо структурным изменениям вириона. Липиды вируса гепатита В и его НВз-антигена образуются путем инвагинации мембран эндоплазматической сети. Вирус герпеса формируется путем почкования через ядерную оболочку, поэтому в его составе есть липиды ядерной оболочки.
3.3. Углеводы вирусов
Углеводный компонент вирусов находится в составе гликопротеидов. Количество Сахаров в составе гликопротеидов может быть достаточно большим, достигая 10—13% от массы вириона. Химическая специфичность их полностью определяется клеточными ферментами, обеспечивающими перенос и присоединение соответствующих сахарных остатков. Обычными сахарными остатками, обнаруживаемыми в вирусных белках, являются фруктоза, сахароза, манноза, галактоза, нейраминовая кислота, глюкозамин. Таким образом, подобно липидам, углеводный компонент определяется клеткой-хозяином, благодаря чему один и тот же вирус, Взращенный в клетках разных видов, может значительно различаться по составу Сахаров в зависимости от специфичности клеточных гликозилтрансфераз.
Углеводный компонент гликопротеидов играет существенную роль в структуре и функции белка. Он является каркасом для локальных участков гликопротеида, обеспечивая сохранение конформации белковой молекулы, и обусловливает защиту молекулы от протеаз. Возможны и другие функции углеводов, пока достоверно не установленные[21] .
Заключение
В итоге работы можем сделать следующие выводы.
Вирусы — это неклеточные формы жизни, различимые только под электронным микроскопом. Это внутриклеточные паразиты. За пределами клетки они не проявляют своих свойств и имеют кристаллическую форму.
Наиболее просто организованные вирусы представляют собой нуклеопротеид, состоящий из нуклеиновой кислоты (ДНК или РНК) и белковой оболочки (капсида). Сложные вирусы могут иметь дополнительную оболочку из липопротеина. Некоторые вирусы (бактериофаги) имеют аппарат транспортировки своего генома в бактерии, после проникновения в клетку капсид остается за пределами клетки. Тело бактериофага имеет сложное строение, оно содержит головку, хвостик (трубку, через которую геном проталкивается в клетку) и хвостовые отростки.
В клетку вирусы могут попасть вместе с пиноцитозными пузырьками или путем погружения части оболочки клетки с приклеившимся к ней вирусом в цитоплазму, а также путем растворения оболочки клетки.
Вирусы вносят в клетку свою генетическую информацию, и клетка начинает производить подобные вирусы.
Внутри клетки начинает синтезироваться ДНК или РНК вируса и образуется множество вирусов. В результате клетка гибнет, и вирусы выходят наружу, заражая новые клетки. Встроенный в геном клетки геном вируса может существовать в таком виде долгое время.
Вирусы вызывают табачную мозаику у растений, оспу, грипп, полиомиелит, гепатит, СПИД у человека. Наибольшую опасность в наше время представляет вирус СПИДа. Он попадает в организм человека при переливании крови, при половых контактах. Этот вирус поражает клетки организма, отвечающие за иммунитет. В результате человек оказывается беззащитным перед инфекционными болезнями и быстро погибает.
Вирусы, благодаря мутированию и способности быстро размножаться внутри клеток, становятся устойчивыми к действию лекарств, и это обстоятельство затрудняет лечение таких вирусных заболеваний, как грипп, гепатит и др.
Список литературы
1. Агол В.И. Биосинтез вирусных нуклеиновых кислот // Молекулярная биология: Структура и биосинтез нуклеиновых кислот / Под ред. А.С.Спирина. М.: 1990.
2. Березов Т.Т., Коровкин Б.Ф. Биологическая химия. – М.: «Медицина», 2004. – 704 с.
3. Борисов Л.Б. Микробиология, иммунология, вирусология. М.: МИА, 2005. – 736с.
4. Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 17-19
5. Вилляреал Л. Вирус: существо или вещество? // В мире науки. 2005. - №3.
6. Вирусология. Под редакцией Филдса Б., Найта Д., тт. 1–3, М.: Медицина, 1989- 492с.
7. Дяченко С. С., Синяк К. М., Дяченко Н. С. Патогенные вирусы человека. – Киев: Здоров'я, 1980. – 448 с.
8. Кобринович Ю.О. Вирусы. [http://zhurnal.lib.ru 13.02.09]
9. Микробиология и иммунология. / Под ред. А.А. Воробьева. – М.: Медицина, 1999. – 464с.
10. Николаев А.Я. Биологическая химия. – М.: МИА, 2004. – 566 с.
11. Поздеев О. К. Медицинская микробиология. – Москва: ГЭОТАР – Мед., 2001.
12. Прозоркина Н. В., Рубашкина Л. А. П 78 Основы микробиологии, вирусологии и иммунологии: Учебное пособие для средних специальных медицинских учебных заведений. — Ростов нД: Феникс, 2002. -416с.
13. Фролов А. Ф., Шевченко Л. Ф., Широбоков В. П. Практическая вирусология. – Киев: Здоровье, 1989.
[1] Прозоркина Н. В., Рубашкина Л. А. П 78 Основы микробиологии, вирусологии и иммунологии: Учебное пособие для средних специальных медицинских учебных заведений. — Ростов нД: Феникс, 2002. -416с.
[2] Вилляреал Л. Вирус: существо или вещество? // В мире науки. 2005. - №3.
[3] Кобринович Ю.О. Вирусы. [http://zhurnal.lib.ru 13.02.09]
[4] Агол В.И. Биосинтез вирусных нуклеиновых кислот // Молекулярная биология: Структура и биосинтез нуклеиновых кислот / Под ред. А.С.Спирина. М.: 1990. С. 260-333.
[5] Микробиология и иммунология. / Под ред. А.А. Воробьева. – М.: Медицина, 1999. – 464с. С. 44-45.
[6] Березов Т.Т., Коровкин Б.Ф. Биологическая химия. – М.: «Медицина», 2004. – 704 с.
[7] Николаев А.Я. Биологическая химия. – М.: МИА, 2004. – 566 с.
[8] Фролов А. Ф., Шевченко Л. Ф., Широбоков В. П. Практическая вирусология. – Киев: Здоровье, 1989. с. 217 – 225.
[9] Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 17-19
[10] Вирусология. Под редакцией Филдса Б., Найта Д., тт. 1–3, М.: Медицина, 1989- 492с.
[11] Поздеев О. К. Медицинская микробиология. – Москва: ГЭОТАР – Мед., 2001. С. 459 – 465.
[12] Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 17-19.
[13] Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 19-22.
[14] Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 19-22.
[15] Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 19-22.
[16] Дяченко С. С., Синяк К. М., Дяченко Н. С. Патогенные вирусы человека. – Киев: Здоров'я, 1980. – 448 с.
[17] Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 22-27
[18] Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 22-27.
[19] Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 28
[20] Борисов Л.Б. Микробиология, иммунология, вирусология. М.: МИА, 2005. – 736с., С. 603.
[21] Букринская А.Г. Вирусология. – М.: Медицина, 1986. – 336с. С. 29-30.