| Скачать .docx | Скачать .pdf |
Реферат: Стебель
П.А. Кошель
Стебли лозы и стволы деревьев, служившие человеку поделочным материалом, всегда были предметом его самого пристального внимания и изучения. Неудивительно поэтому, что уже на первых шагах развития ботанических знаний, в древнегреческой науке, мы встречаем довольно обстоятельные сведения о строении стебля.
Теофраст в структуре стебля растений различает кору, древесину и сердцевину. Пытаясь разобраться в деталях каждого из этих трех структурных элементов стебля, Теофраст приходит к выводу, что они состоят из волокон, жил и мяса. Волокна и жилы, по Теофрасту, представляют собой в сущности одно и то же, но волокнами или нервами он называл тонкие волокноподобные пучки, а жилами – более крупные тяжи (по-видимому, сосудистые пучки).
«Волокна и жилы, – говорит Теофраст, – расщепляются только вдоль и не иначе, между тем как мясо растений можно легко делить во всех направлениях подобно кому земли».
В этом определении мы можем видеть начало той старой анатомической классификации растительных тканей, которая и теперь еще сохранила свои следы в терминах «прозенхима» и «паренхима».
Теофраст утверждал, что древесина состоит преимущественно из «волокон», «жил» и влаги, сердцевина – из «мяса» и сока, а кора – из «волокон», «мяса» и сока.
Теофрасту были уже известны различия внутреннего строения тех двух больших групп растений, которые мы теперь называем однодольными и двудольными. Он задумывался и над вопросом о причинах образования так называемых годичных колец в древесине крупных стволов строевого леса.

Годичные кольца и строение древесины
Творения Теофраста на протяжении многих веков последующей истории человечества были единственным источником сведений, которые наука могла предложить по данному вопросу. Только в XVI в. нашей эры Чезальпино попытался дополнить картину строения стебля, нарисованную Теофрастом, собственными наблюдениями и соображениями.
Нельзя сказать, чтобы Чезальпино внес много нового в учение о строении стебля по сравнению с Теофрастом. Рассуждения Чезальпино содержат чересчур смелые аналогии в строении тел растений и животных, но в них он высказывает одну правильную мысль: волокна, или нервы, и жилы растений, он считает мельчайшими полыми трубочками, служащими для проведения влаги и обладающими столь малыми просветами полостей, что заметить их глазом невозможно.
Он произвольно приравнивает волокна стебля к венам в теле человека и животных. К этому сравнению он приходит, наблюдая выделение млечного сока из стебля некоторых растений. Вот рассуждения Чезальпино по этому вопросу.
«Что вены даны растениям, видно хотя бы по тем проходам, которые выделяют млечный сок у молочая и фикуса. Если их срезать, выделяется много сока, который течет, как кровь, из разрезанного мяса животного. Много этого сока заключается и в виноградной лозе. Вследствие крайней узости этих каналов для движения сока их невозможно различить глазом...
...На всех стеблях и корнях можно бывает видеть нечто похожее на длинные нервные волокна. Нужно думать, что эти волокна представляют собой каналы для проведения пищи; по своему положению они соответствуют венам животных. Правда, у растений нет ни одной стволовой вены (подобной полой вене животных), но зато у них имеется множество тонких каналов, идущих из корня в сердце, а затем оттуда восходящих в стебель».
Только через 100 лет после Чезальпино наука, получившая в свое распоряжение первые несовершенные еще микроскопы, смогла представить более обстоятельную картину строения стебля благодаря работам одного из отцов микроскопической анатомии растений – Марчелло Мальпиги (1628–1694).
Мальпиги открывает в стебле подлинные трубки, или сосуды, снабженные к тому же внутри спиральной лентой, как бы препятствующей спаданию их стенок. Современник Мальпиги, английский врач Неемия Грю, подтверждает наблюдения своего итальянского коллеги и дает описание микроскопической структуры древесины и коры стебля, представляющихся ему сплетением горизонтальных и вертикальных волокон наподобие тонкого кружева.
Мальпиги, занимавшийся и анатомией животных, оказался так же, как и Чезальпино, не вполне свободным от пристрастия к проведению рискованных аналогий в строении тел животных и растений. Он считал открытые им в стебле растений спиральные сосуды вполне аналогичными дыхательным трубкам, или трахеям, в теле насекомых, поэтому он предложил назвать открытые им спиральные сосуды трахеями. Он считал, что эти трахеи в теле растения также выполняют дыхательную функцию. Любопытны рассуждения самого Мальпиги по этому вопросу.
«Легкие у животных, – говорит Мальпиги, – тем больше и тем сильнее развиты, чем организация животного примитивнее. От человека, который имеет пару сравнительно маленьких легких, можно провести линию сравнения до насекомых, у которых аппарат дыхательных трубочек распространен по всему телу. ...У растений, которые, надо полагать, относятся к самым низшим организмам, естественно ожидать, что трахеи должны быть развиты в еще большей степени, в таком размере, что они проходят через все мельчайшие части их тела за исключением коры. Если наземные животные черпают воздух для дыхания из атмосферы, а рыбы – из воды, то растения получают свой материал для дыхания из земли или, правильнее, из воды и воздуха, которые смешаны с землей».
Доказательство правильности своих мыслей Мальпиги видит в том, что спиральные трубки в корне крупнее и их больше. Это ошибочное учение о роли спиральных сосудов как органов дыхания растений, а не проведения воды, держалось в науке более 100 лет и отразилось в самом названии их, сохранившемся до наших дней, – трахеи, трахеиды.
В том же XVIII в., кроме Мальпиги и Грю, вопросами строения стебля интересовался Антони ван Левенгук. Он при помощи своих удивительных по тонкости конструкций миниатюрных луп обнаружил, что в трубках древесины бывают утолщения не только спиральной, но и других форм.
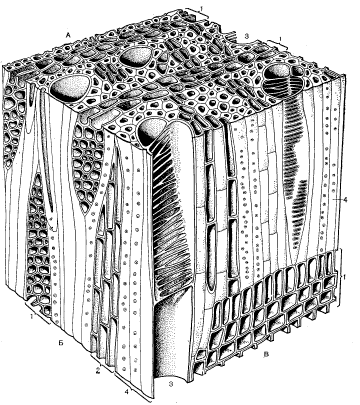
Трехмерная блок-диаграмма древесины дегенерии фиджийской:
А – поперечный разрез; Б – тангентальный разрез; В – радиальный разрез:
1 – древесинные лучи; 2 – древесинная паренхима; 3 – сосуд; 4 – волокна
Бернгарди (1774–1850) подробно описал строение так называемых кольчатых сосудов древесины. Длинные пучки сосудов, описанные Мальпиги и Левенгуком (наряду с отмеченным ими же наличием в теле растений коротких или округлых клеток или пузырьков), долгое время занимали в системе тканей растения какое-то обособленное положение.
Первые попытки вывести и форму сосуда из формы пузырька, или клетки, как основной формы внутренних структурных элементов растений, принадлежат Г.Тревиранусу (1776–1837). Ему удалось привести первые доказательства клеточного происхождения сосудов. Тревиранус указывал на то, что в тканях совсем молодых листьев еще нет вполне выраженных сосудов, но со временем в клетках, расположенных продольным рядом, поперечные перегородки исчезают, и из ряда клеток образуется один сосуд.
Клеточное происхождение сосудов было окончательно доказано Г.Молем, подробно проследившим процесс развития сосудов в различных органах и доказавшим, что сосуд есть продукт срастания отдельных клеток. Современную систему классификации элементов древесины с подразделением их на паренхиматические, механические и сосудистые дал в 60-х гг. XIX в. прусский школьный учитель Санио. Он предпринял обширные исследования строения древесины различных групп древесных растений, заложив основы метода сравнительно-анатомического изучения стеблей двудольных и голосеменных. Ему же мы обязаны детальным и обстоятельным изучением и описанием устройства окаймленных пор в клетках древесины хвойных, первое описание которых дано было ранее Шахтом.
Бинокулярный микроскоп, который использовали в 1685 г.
Теодор Гартиг (1805–1880) занимался изучением флоэмы. Он открыл так называемые ситовидные трубки, по которым в стебле передвигается пластический материал, произведенный в листьях. Гартиг обнаружил, что эти трубки состоят из цепочки клеток, поперечные перегородки между которыми снабжены сквозными отверстиями.
На основе этого открытия Гартига К.Негели затем разработал целую теорию функций ситовидных трубок как органов проведения трудно диффундирующих пластических веществ по стеблю растений. Теория эта в ее основных чертах сохраняет свое значение и в настоящее время. Что касается распределения сосудистых элементов в стебле растений, то еще в самом начале XIX в. Д.Мольденгауэр (1766–1827) подметил, что в стеблях однодольных растений сосудистые элементы соединяются с группами паренхимных клеток и волокон в особые тяжи, получившие название сосудисто-волокнистых, или, просто, сосудистых пучков. На стебель двудольного Мольденгауэр смотрел как на продукт слияния таких сосудистых пучков в одно кольцо. Наружная часть этих слившихся в одно кольцо пучков, содержащая ситовидные трубки, образовала наружную цилиндрическую трубку флоэмы, а внутренняя часть с водоносными сосудами – внутренний цилиндр древесины.
Каждый из многих сосудистых пучков, разбросанных в толще стебля однодольного растения, как выяснил Мольденгауэр, в периферической части состоит из нескольких слоев толстостенных клеток с косыми щелевидными порами на внутренних стенках. Эти клетки, примыкающие и к лубяной, и к древесинной части пучка, вызвали среди анатомов середины XIX в. споры о том, куда их следует отнести – к элементам флоэмы или ксилемы. Затруднение это в 1874 г. разрешил Швенденер, предложивший выделить эти элементы (встречающиеся к тому же во множестве и вне пучков) в самостоятельную группу механических элементов, или стереид, т.к. они служат преимущественно для сообщения надлежащей крепости данному органу, образуя как бы скелет растения.
Так были открыты и описаны главнейшие микроструктурные элементы стебля. Однако первые ботаники-исследователи не были специалистами, узко ограничивавшими свои исследовательские интересы вопросами микроскопической анатомии растений. Своей целью они считали также и раскрытие физиологических функций изучавшихся ими структурных образований. Как ни наивны были самые первые попытки научных исследователей в этой области, мы все же должны упомянуть о них.
В очерке, посвященном изучению корня, мы уже говорили о теории Чезальпино (конец XVI в.), утверждавшего, что растение всасывает своими корнями влагу так же, как фитиль светильника тянет масло к огню. Передвижение воды по стеблю Чезальпино объяснял тягой соков к началу внутренней теплоты растений. Этому движению соков в побегах, по мнению Чезальпино, способствует свойственное растениям тепло, которое поглощает поднятую влагу и вызывает новое всасывание. Кроме внутренней теплоты, подъем соков по стеблю объясняется, по Чезальпино, и жаром летних лучей солнца.
«Так как тепло гонит влагу вверх, то большинство растений лучше всего растет и приносит плоды весной и летом, потому что тогда от внешнего тепла увеличивается приток влаги. Зимой пламя жизни растений угасает, загнанное глубоко внутрь их тела, при угасании жизни влага больше не распространяется в растении. Органы передвижения влаги засыхают и погибают или твердеют и становятся деревянистыми, как это бывает в старости».
Здесь, наряду с фантастическим утверждением о существовании какой-то движущей силы – «внутренней теплоты растений», – мы видим и зачаток правильных представлений о действительной зависимости подъема воды в сосудах стебля от инсоляции, вызывающей, как мы знаем сейчас, усиление процесса транспирации.
Один из «отцов анатомии растений», Неемия Грю, для объяснения процесса поднятия воды по стеблю пытался проводить аналогию между работой сосудистой системы растений и работой кровеносной системы животных.
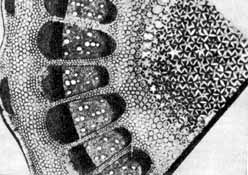
Рисунок поперечного сечения стебля чертополоха, сделанный Грю в 1672 г. для книги «Анатомия растений»
Грю считал, что вода в стебле передвигается вверх благодаря ритмическим сокращениям (пульсации) паренхимы, окружающей сосуды. Сдавливая сосуды, паренхима, по мнению Грю, должна выжимать воду все выше и выше.
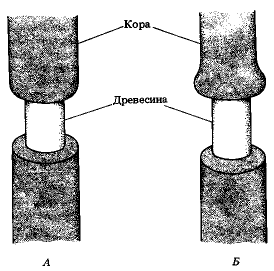
Опыт Мальпиги со снятием кольцеобразного куска коры со стебля (А). Набухание ткани над кольцом (Б)
Мальпиги, как мы уже знаем, вообще не считал открытые им спиральные сосуды за водоносные пути стебля, а называл их трахеями, приписывая им, как и трахеям насекомых, дыхательную функцию. При этом Мальпиги, естественно, должен был задуматься над вопросом: по какой же части ствола совершается восходящий ток влаги из корней к листьям и плодам? Прежде всего его внимание привлекла кора дерева. «Не может ли совершаться восхождение влаги по коре?» – спрашивал себя Мальпиги и для решения этого вопроса поставил первый опыт с кольцевой вырезкой на коре дерева (1679). Он обнаружил, что части растения, расположенные выше такой кольцевой вырезки, не погибают, а продолжают снабжаться водой, из чего он заключил, что ток воды передается листьям не по коре, а по каким-то элементам древесины. Продолжая свои наблюдения над окольцованными деревьями, он с удивлением заметил, что через год части деревьев, расположенные выше кольцевых вырезок коры, разрослись сильнее обыкновенного в толщину, а плодовые деревья принесли большее против обычного количество плодов. При этом особенно сильно разрастался ствол у верхнего края вырезки – здесь образовался даже кольцеобразный наплыв. Совершенно обратное замечалось в части ствола ниже вырезки. Со времени вырезывания кольца коры здесь прекращался рост дерева в толщину. Из всех этих наблюдений Мальпиги должен был сделать вывод о том, что кора служит, главным образом, для передвижения пластических веществ вниз, а ток воды с питательными солями почвы поднимается какими-то путями по древесине.
Таким образом, было положено начало учению о существовании в стебле растений двух токов растворенных веществ: восходящего по древесине и нисходящего по лубу. Обстоятельно разработано это положение было в целом ряде опытов Гельса, описанных нами ранее в очерке, посвященном строению и работе корня. Гельсу удалось открыть и два так называемых концевых двигателя восхоящего тока: корневое давление и присасывающее действие испаряющей работы листьев. Некоторые натуралисты того времени полагали, что по аналогии с венами животных сосуды древесины должны иметь на своей внутренней поверхности клапаны, препятствующие обратному току воды. Гельс с полной очевидностью доказал отсутствие в сосудах древесины каких бы то ни было клапанов и возможность передвижения в них воды в обоих направлениях. Это положение было доказано следующим остроумным опытом. Гельс привил молодое дерево ветвями к двум соседним деревьям и затем срезал привитое дерево под корень, так что оно утратило питание от собственных корней и осталось висеть как бы подвешенное к ветвям соседних деревьев. Срезанное дерево оставалось живым и развивало листья, несмотря на то, что оно получало теперь воду через ветви соседних деревьев.
Схематическое изображение подсолнечника, который использовал Гельс в работах по изучению движения воды в растениях
Гельс доказал также, что в стволе дерева возможно передвижение воды не только снизу вверх, но и горизонтально (хотя в этом случае наблюдается сильное замедление тока воды). Это положение было доказано на следующем опыте. Гельс сделал на отрезанной ветви дуба две глубокие диаметрально противоположные вырезки, доходившие до самой сердцевины на расстоянии 4 дюймов одна от другой; ветвь, погруженная нижним концом в воду, осталась свежей, несмотря на значительное испарение воды. Подобный же результат был получен им при вырезывании из ствола четырех накрест поставленных одна над другой вырезок, доходивших до самой сердцевины. Из своих опытов Гельс сделал вывод, что хотя, как правило, восходящий ток идет и древесине деревьев по прямой линии, но он может осуществляться и в других направлениях (очевидно, благодаря наличию сообщения смежных сосудов между собой).
После Гельса изучением явлений восходящего и нисходящего токов в стебле растений занимался Т.Найт. В 1801 г. он применил для обнаружения проводящих тканей древесины окрашенные растворы и выяснил, что краска вместе с водой из стакана поднимается по срезанному стеблю растения только через сосуды древесины, в то время как сосуды коры, видимо, не принимают в этом подъеме жидкости никакого участия, так как остаются неокрашенными.
Опыты Найта наглядно подтвердили учение о восходящем и нисходящем токах в стебле растений.
Теория пассивного водоносного ложа с двумя концевыми двигателями, основанная на опытах Гельса, впервые была четко сформулирована в 1837 г. А.Дютроше (1776–1847). Однако вскоре этой теории был предъявлен ряд серьезных возражений, основанных на учете количественных отношений. Прежде всего указывалось, что корневое давление достигает величины не больше двух атмосфер; следовательно, нижний концевой двигатель мог бы поднимать воду до верхушки стебля только у небольших травянистых растений. Большей оказалась величина сосущей силы клеток листовой паренхимы. Но если принять во внимание высоту некоторых гигантов растительного мира, например калифорнийской секвойи (100 м) или австралийских эвкалиптов (150 м), то для подъема воды силы указанных двух двигателей представлялись недостаточными. В подтверждение этих сомнений указывали также на большое сопротивление, которое приходится преодолевать току воды при прохождении через сосуды древесины весьма высоких деревьев.
Чтобы обойти эти возражения, некоторые авторы предлагали учитывать капиллярные свойства древесины, но вычисления показали, что в древесине хвойных капиллярное поднятие воды не может превышать 2–3 м. Тогда от капиллярных трубок воображение теоретиков перекинулось к межмолекулярным ходам, к идее движения воды в самой толще стенок сосудов.
Разработанная на этих принципах в 1870-х годах так называемая имбибиционная теория Сакса ненадолго привлекла к себе внимание ученых Запада и была высмеяна Тимирязевым, заявившим, что эта теория «...приписывает клеточной стенке физические свойства, не встречающиеся ни в каком другом теле».
Тимирязев считал эту теорию практически неверной и теоретически недопустимой.
Как всегда в моменты временных теоретических затруднений, после этого стали одна за другой нарождаться виталистические гипотезы под именем учений о промежуточных двигателях восходящего тока. Эти теории выдвигали идею активного участия живых клеток стебля в проведении воды. Вместо определенной теоретической схемы они давали ряд неясных и туманных указаний на внутреннюю регуляцию процессов сокодвижения в стебле. Еще большую путаницу в понимание данного вопроса внесло открытие так называемых жаменовских цепочек, т.е. целого ряда пузырьков воздуха, которыми будто бы всегда внутри сосудов ток поднимающейся воды разбит на мелкие части. За эти жаменовские цепочки ухватились представители обоих направлений. Материалисты видели в менисках пузырьков давно искомую «точку опоры» для столба воды, поднимающегося на большую высоту; виталисты же, наоборот, говорили о большом сопротивлении, которое эта система пузырьков воздуха представляет собой для поднимающегося тока жидкости.
Тщательные наблюдения внесли некоторую ясность, констатировав, что эти пузырьки, представляющие собой в сосудах далеко не постоянное, а скорее исключительное явление, в сущности не препятствуют непрерывному току воды.
Первым толчком к разрешению вопроса послужили опыты Бема, предпринятые им в 1840 г. Бем исследовал величину присасывающей силы испаряющих воду листьев. Он срезал над водой ветви разных растений и укреплял их в длинной стеклянной трубке, наполненной водой и погруженной нижним концом в воду. По мере испарения воды листьями ветка всасывала воду из трубки и вызывала поднятие столбика ртути. Высота поднятия ртути служила мерой «силы сосания» листьев. В некоторых из этих своих опытов Бем обнаружил парадоксальное на первый взгляд явление: ртуть поднималась иногда на высоту, значительно большую той, которая имелась в барометрической трубке (в которой, как известно, под столбиком ртути нет никакого давления воздуха).
Для понимания причин этого явления надо иметь в виду следующее: между сосущим концом ветки и ртутью находится столб воды. Для остановки ртути на какой-нибудь высоте необходимо, чтобы произошел разрыв этого столба воды, но сила сцепления частиц воды оказывается поистине громадной. Отсюда следует, что присасывание испаряющих листьев может удерживать и тянуть вверх тонкие водные нити громадной длины. На базе этих данных Диксон и Джоли в 1895 г. сформулировали новую теорию в учении о двигателях восходящего тока. Согласно этой теории, главнейшим и основным двигателем восходящего тока (особенно в период сильной транспирации – летом) является «сосание кроны», которое в связи с силой сцепления частиц воды распространяется до самого корня, независимо от высоты растения. Эта теория предполагает, что в растении от адсорбирующей ткани корня до испаряющей поверхности паренхимы листьев тянутся тончайшие, но непрерывные, прочные на разрыв нити воды.