| Скачать .docx | Скачать .pdf |
Реферат: Морфологическая изменчивость у колониального гидроида dynamena pumila и классификация обнаруженных морфотипов книдарии
. Современное состояние и перспективы исследований. Ii/спб.. 1995 (труды зоологического института ран. Т. 261)
H. Н. Марфенин, Р. Я. Маргулис, Е. М. Майер
Разработан метод анализа морфологической изменчивости популяции колониального гидроида Dynamena pumila на основе учета отклонений от нормы формы перисарка стереотипных междоузлии побегов, называемых морфологическими аномалиями или морфотипами. По результатам обработки более 200 000 междоузлии составлена классификация морфологических аномалий, включающая 7 разделов и 47 разновидностей. В статье представлены описания и рисунки всех морфотипов. Материал собран на Белом и Баренцевом морях в 1991—1993 гг.. а также 100 лет тому назад (коллекции Зоологического института РАН). Некоторые морфотипы выглядят как полноценные резервные фенотипы вида. Для большинства морфотипов приведены объяснения возможных причин их образования.
Из всех гидроидов пока что только для одного вида описана изменчивость строения стереотипных частей колонии — междоузлии побегов (Марфенин, 1975, 1988), Этот вид—Dynarnena pumiia (L.) из высокоорганизованного семейства Sertulariidae отряда Thecaphora — обладает в норме двурядным супротивным расположением гидрантов на стволе побега (рис. 1). В процессе роста побега очередная пара гидрантов формируется на его верхушке. Хитиновый перисарк, выделяемый здесь, принимает во время морфогенеза верхушки соответствующую форму, имеющую видоспецифический характер. Морфогенез побега протекает циклично, так что к концу образования очередной пары гидрантов воссоздаются исходные условия для формирования следующей пары (рис. 2). Каждый стереотипный участок побега, включающий пару гидрантов и отрезок ствола, ясно отграничен от следующего подобием перетяжки, что позволяет выделять в побеге соответствующие междоузлия.
Таким образом, типичный для Dynamena pumiia морфогенез повторяется в каждой колонии множество раз, что позволяет изучать фенотипическую изменчивость даже в пределах отдельно взятого генетически однородного организма, каковым на самом деле и является колония у гидроидов (Марфенин, 1993). Подобную задачу можно ставить и на других так называемых модульных организмах, вроде растений и микромицетов, или же на совокупности клеток одного организма, однако до сих пор такие исследования единичны.
Возможно, это обстоятельство объясняется излишней вариабельностью строения модулей (листьев, побегов),т.е. недостаточной стабильностью их канонической формы, что затрудняет выделение четких критериев для исследования внутривидовой изменчивости. Вторая возможная причина связана с недостаточностью проявления вариабельности строения модулей у многих видов. Например, у гидроидов из семейства Campanulariidae нам не приходилось встречать обращающих на себя внимание морфологических отклонений от нормы в строении частей колонии.
Dynamena pumila в этом отношении значительно отличается от других гидроидов, так как ненормальные по своему внешнему виду междоузлия встречаются в ее колониях довольно часто. По существу у D. pumila мы встречаем разнообразие морфотипов. представленное как доминирующей формой междоузлия, принятой за норму, свойственную данному виду, так и значительно реже встречающимися другими формами. В большинстве случаев они выглядят уродливо и поэтому первоначально были названы аномалиями формы побега (Марфенин, 1975). Однако среди аномалий встречаются морфологически и функционально вполне полноценные формы, которые могут последовательно и относительно устойчиво воспроизводиться в побеге. И те, и другие отклонения от канонической формы обладают постоянными индивидуальными чертами строения, отличными друг от друга, т. е. присущи виду, поэтому правильнее их в общем случае называть морфотипами, хотя для многих явно уродливых образований мы оставляем название морфологические аномалии.
Полноценному изучению феномена внутриколониальной морфологической изменчивости Dynamena pumila должны предшествовать описание и систематизация морфотипов, чему и посвящена настоящая статья. Если первоначально были описаны 27 морфотипов неканонического строения междоузлии побега D. pumila (Марфенин, 1975), то после специального исследования тысяч побегов нами выделено множество аномалий, которые сведены к 47 морфотипам.
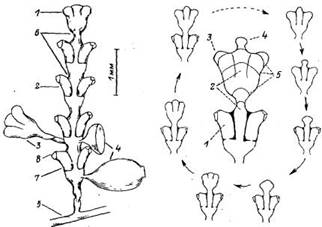
Рис. 1. Схема строения нормального побега Dynamena pumila:
1 — верхушка роста ствола побега; 2 — ствол побега; 3 — боковая ветвь побега (7 порядка); 4 — гонотенн на разных стадиях развития; 5 — столон; б — перетяжки побега (границы неждоузлий; 7— гидротека (нижняя часть гидротеки — диафрагма); 8—устье гидротеки
Рис. 2. Морфогенетический цикл образования междоузлия у побега Dynaтепа pumila.
I — нормальная гидротека сформировавшегося гидранта; 2 — ствол побега; 3 — формирующийся гидрант; 4 — верхушка роста на ранней стадии морфогенеза; 5—последовательные характерные стадии развития верхушки
Материал и методика
К настоящему времени удалось обстоятельно изучить две популяции Dynamena pumila: одна из Ругозерской и Кислой губ Кандалакшского залива Белого моря, а другая из губы Дальне-зеленецкой Баренцева моря. Пробы были собраны летом 1991 и 1992 гг. на Белом море нами, а на Баренцевом море по нашей просьбе сотрудницей ММБИ К. Н. Пантелеевой (за что приносим ей искреннюю благодарность).
Дополнительно был использован музейный материал из коллекции Зоологического института РАН, собранный в Белом море (о-в Сидоров) в 1895 г., а в Баренцевом море в 1893 г. (Екатерининская гавань) и в 1899 г. (о-в Седоватый). Сравнительный анализ изменения встречаемости морфотипов D. pumila за 100 лет будет рассмотрен в отдельной статье. В данном исследовании мы хотели составить как можно более полное представление о спектре присущих виду морфотипов по всему имеющемуся у нас материалу.
Для исследования отбирали малообросшие колонии с субстратом. Побеги отрезали у основания и фиксировали, или же фиксировали колонии вместе с субстратом. Следует иметь в виду, что в колониях могут быть уродливые крошечные побеги, которые при отборе более крупных остаются незамеченными и неучтенными. При обработке материала побеги просматривали под бинокуляром. В журнале регистрации записывали: 1) порядковый номер; 2) число междоузлии в стволовой части побега; 3) число междоузлии в каждой боковой ветви побега (сначала ветви 1 порядка, затем 11 порядка и т. д.); 4) суммарное число междоузлии и побеге; 5) номера морфотипов по описанной ранее классификации (Марфенин, 1975) или же номер рисунка, не попадающего эод прежнюю классификацию морфотипа; б) месторасположение лорфотипа (число междоузлии до аномалии), считая от верхушки побега. Все побеги с отклонениями от канонического морфотипа сохраняли в коллекции.
Правильнее всего оценивать выборки по суммарному числу обследованных междоузлии и лишь затем — по числу побегов. Зсего было исследовано более 200 тыс. междоузлии беломорской юпуляции и более 110 тыс. междоузлии баренцевоморской популяции D. pumila. Соответственно, более 8000 и 5000 побегов.
В конечном результате по каждой выборке можно было указать совокупную долю (%) всех неканонических морфотипов и встречаемость (%) каждого из них. В данной статье мы будем считать, ito морфотип встречается: а) очень редко, если доля его находок во всей совокупной выборке составляла менее 0.002%, 5) редко: от 0.01 до 0.002%, в) периодически: 0.05— 3.011%; г) часто: 0.1—0.051%; д) очень часто:более 0.1%
Регистрация расположения морфотипов в колонии не дала каких-либо интересных результатов. Аномалии распределялись в пределах побега достаточно равномерно.
Систематизация морфотипов представляет собой самостоятельную задачу, которую мы обсудим ниже, после основного раздела, содержащего их описания. Мы остановились на самом простом и достаточно формальном подходе: морфотипы условно разделены по группам в соответствии с их местоположением в колонии. Всего получилось 7 групп.
Цифровые обозначения присваиваются морфотипам по следующему порядку. Первая (римская) цифра обозначает номер группы. Через тире указывается арабскими цифрами присваиваемый порядковый номер морфотипа. Близкие по своему строению аномалии получают номера в пределах одного десятка. Пропущенные в пределах десятка номера резервируются для будущих находок и одновременно маркируют границы подгрупп. В том случае, когда аномалия была описана (Марфенин, 1975), в скобках приводится ее прежний номер. Если таковой отсутствует, морфотип описан впервые.
Описание различных отклонений от нормального морфотипа dynamena puмila
Группа I: Отклонения в морфогенезе ствола побега без остановки его дальнейшего роста (рис. 3)
Аномалии выражаются в необычном взаимном расположении междоузлии, гидрантов, а также необычной форме междоузлии. Форма гидротек остается нормальной.
I—1 (1), Изгиб ствола без деформации междоузлия может происходить в самые различные стороны относительно продольной оси побега. Наибольший угол отклонения равен 90°. Местом изгиба является исключительно перетяжка. Возникает после приостановки роста побега в том случае, если при начале очередного периода роста верхушка ценосарка растворяет хитиновую теку не апикально, а сбоку. Кроме того, аномалия возникает в том случае, если растущий побег встречает на своем пути преграду. Встречается периодически.
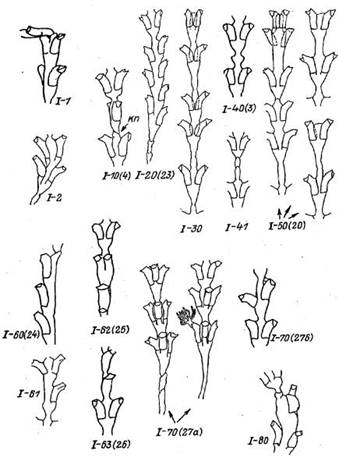
Рис. 3. Отклонения в морфогенезе ствола побега Dynamena pumUa без остановки его дальнейшего роста (группа I)
кп— косая перетяжка
1—2. Изгиб ствола с деформацией междоузлия. Изгиб происходит в процессе роста верхушки побега и в пределах его фронтальной плоскости. С приостановкой роста изгиб ствола не связан. Причиной может быть несоответствие скоростей роста двух сторон верхушки. Встречается редко.
1—10(4). Поворот плоскости колонии вокруг собственной оси. Местом поворота является тонкая перетяжка. Поворот может быть на самый различный угол, вплоть до 90°. Возникает после приостановки роста. Часто сопровождается формированием косой перетяжки (рис. 3, 10, к.п) после начала следующего периода роста. В норме косая перетяжка образуется только на зачатке основного побега при его образовании на гидроризе. Встречается периодически.
1—20 (23). Поочередное расположение гидрантов. Морфотип представляет собой смещение гидрантов одного междоузлия по отношению друг к другу по вертикали. Чаще всего первоначальное смещение незначительно, однако. если несколько пар гидрантов подряд отделены друг от друга толстыми перетяжками, то в каждой последовательной паре смещение увеличивается. После образования тонкой перетяжки смещение исчезает. Самое распространенное отклонение от канонического строения Dynamena pumila. Среди всех аномалий на ее долю может приходиться от 1/3 до 1/2 случаев, что составляет 0.6—0.7% от всех нормальных междоузлии.
I—30. Смещение супротивно расположенных гидрантов навстречу друг другу. В норме гидротеки междоузлия расположены не точно в одной плоскости со стволом побега, а примерно под углом 150—165° друг по отношению к другу. Еще большее сближение гидротек в междоузлии по отношению друг к другу представляет собой одну из аномалий. Ствол побега в таком случае проходит позади сближенных гидротек. Встречается редко.
1—40(3). Утолщение на стволе. Утолщение цено-сарка между двумя парами гидрантов. Диаметр утолщения равен 1.25—2.0 диаметра перетяжки. Обычно возникает 1 утолщение, но может быть и 2. Утолщение возникает в случае приостановки роста верхушки (еще не расчленившейся в верхней части на 3 зачатка) с последующим ростом только центральной части верхушки. Встречается очень часто.
1—41. Утолщения на стволе, соответствующие по форме ножке побега. Ножка побега имеет характерное четковидное строение в виде 2—3 слабых вздутий перисарка, разделенных косыми перетяжками и следующих друг за другом. Появление такой структуры между парами гидрантов аномально. Встречается периодически.
1—50(20). Цилиндрическая длинная перетяжка между парами гидротек. В норме перисар-<альная трубка ствола на участке между соседними парами гидротек сначала сужается в направлении роста, а затем сразу расширяется (см. рис. 2). Так образуется перетяжка между очередными парами гидротек (или междоузлиями). В данном случае ствол на некотором протяжении остается неизменного сечения, формируя удлиненную цилиндрическую перетяжку, что необычно для нормального междоузлия побега. Встречается периодически.
I—60(24). Однорядное расположение гидрантов. Аномалия представляет собой оригинальную форму бокового или, реже, основного побега, которая состоит из одного ряда гидрантов и ценосарка. Гидротеки поочередно развернуты своими устьями налево и направо. Обычно аномалия повторяется на протяжении нескольких междоузлии. Между междоузлиями перетяжек не бывает. После образования перетяжки побег растет нормально, образуя двухрядное расположение гидротек. Данная аномалия совершенно определенно представляет одну из редко встречающихся (недоминирующих) нормаль.ных форм D. pumila. "Однорядное" строение характеризуется морфологической полноценностью, повторяемостью (т. е. воспроизводится в циклическом морфогенезе побега), жизнеспособностью. Ее образование не вызывает никаких нарушений ни в морфогенезе гидрантов, ни в росте побега. В то же время морфогенез побега принципиально отличен от обычного, так как верхушка раз за разом устойчиво расщепляется в процессе роста на 2 зачатка вместо 3, как должно быть в норме. В деталях такое строение весьма напоминает расположение гидрантов у Hydrallmania falcata. К этой аномалии полностью применимо понятие морфотип. Встречается часто.
1—61. Одно рядность вследствие формирования лишь одного гидранта в междоузлии. Аномалия возникает по иным причинам, чем предыдущая. При формировании междоузлия сохраняется обычный для D. pumila морфогенез, однако один из двух симметричных зачатков гидрантов почему-то не развивается. Возможно, подобная аномалия — следствие приостановки роста верхушки с последующим возобновлением роста только зачатков ствола и одного из двух гидрантов. Встречается редко.
I—62 (25). Переход однорядного расположения гидрантов в двухрядное. Аномалия представляет собой постепенный переход от однорядного расположения гидрантов в двухрядное по мере роста побега. Сначала образуется гидрант больших, чем обычно, размеров, затем в следующем междоузлии раздвоенный гидрант, в следующем — два сближенных гидранта и, наконец, два нормальных, супротивно расположенных гидранта. Указанный переход может реализовываться и быстрее, если одна из промежуточных стадий выпадает. Встречается редко.
I—63 (26). Переход двухрядного расположения гидрантов в однорядное. Аномалия аналогична предыдущей, только порядок перехода противоположный. Встречается очень редко. В исследованной нами огромной выборке не была отмечена ни разу, хотя ранее была однажды отмечена (Марфенин, 1975).
I—70(27а). Трехрядное расположение гидрантов. Аномалия представляет собой правильное трехрядное расположение гидрантов, повторяющееся, чаще всего, в последующих междоузлиях. Иногда расположение гидрантов оказывается не столь правильным [см. I—70(276)], что выражается в уменьшении линии контакта прилежащей стороны одной из гид-ротек с ценосарком. Механизм аномалии неясен. По своей полноценности, повторяемости и жизнеспособности эту аномалию также можно классифицировать как редкий морфотип D. ритilа. Встречается редко.
I—80. Уродливое разрастание ценосарка побега. Разнообразные морфогенети чески неустойчивые уродливые утолщения перисарка ствола, рядом с которыми могут быть уродливые гидротеки. Встречается редко.
Группа II/ Отклонения в морфогенезе верхушки побега, препятствующие ее дальнейшему росту (рис. 4)
II—1(12). На верхушке побега гидрант. Гидрант ненормален по форме, что выражается либо в его больших размерах, либо в необычной искривленности гидротеки. В результате этой аномалии дальнейший рост побега становится невозможен. Встречается редко.
II—2. Гидрант вместо побега. Единичный гидрант на столоне вместо побега. Имеет характерную для побега ножку с несколькими косыми перетяжками. Встречается очень редко.
II—10(8). Две нормальные по форме, но сросшиеся гидротеки почти или полностью без промежутка между ними. Аномалия свойственна верхней паре гидрантов побега. В крайнем своем выражении аномалия представлена двумя гидрантами, между которыми отсутствует ценосарк. Чаще остается ничтожный просвет, в котором невозможно его нормальное развитие ценосарка. Из экспериментов ясно, что аномалия возникает при голодании колонии, когда перестает расти верхушка, которая находится на поздних стадиях роста. В этом случае гидранты все равно дорастают до конца, хотя все междоузлие образуется меньших, чем обычно” размеров. Встречается редко.
II—11. Две уродливые сросшиеся гидротеки без ценосарка между ними. Аномалия аналогична предыдущей за важным исключением — морфогенез верхушки нарушен, и форма образующихся гидротек уродлива. Встречается очень редко.
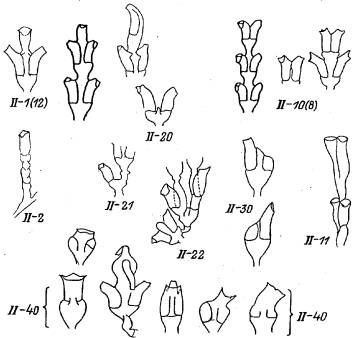
Рис. 4. Отклонения в морфогенезе верхушки побега Dynamena pumiia, препятствующие ее дальнейшему росту (группа II)
II—10. Две независимые гидротеки без участка ценосарка между ними. Очевидно, два гидранта формируются на верхушке побега после остановки ее роста из двух крайних зачатков. Средний зачаток при этом не растет. Встречается очедь редко.
II—21. Гидрант и боковая ветвь из верхушки побега. Аномалия подобна предыдущей, только из одного крайнего зачатка образуется гидрант, вероятно, после остановки роста верхушки, а из другого — боковая ветвь. Средний зачаток не растет. Встречается очень редко.
II—22. Оба зачатка гидрантов дают побеги, а средний зачаток не растет. Аномалия аналогична предыдущей. То, что аномалия на приведенном рисунке расположена в первом междоузлии боковой ветви, а не ствола, принципиального значения не имеет. Специфическая особенность приведенного на рисунке примера в том, что первые междоузлия ветвей, возникших из зачатков гидрантов, однорядные. Встречается очень редко.
II—30. Один боковой гидрант на верхушке побега. После прекращения роста верхушки побега один из зачатков гидрантов продолжает развиваться, тогда как другой и зачаток ствола остаются неразвитыми. Встречается очень редко.
11—40. Уродливая неполноценная верхушка побега. В эту категорию могут быть зачислены разнообразные примеры крайнего нарушения морфогенеза верхушки, приводящие к остановке ее роста и препятствующие возвращению к нормальному циклическому морфогенезу. Подобные аномалии встречаются очень редко.
Группа III: Ювенильные уродливые побеги (рис. 5)
Особая группа аномалий, проявляющихся в момент образования побега на столоне, когда первое же междоузлие выглядит уродливо, а сам побег теряет способность расти дальше. Такие крошечные уродцы ранее не попадали в поле нашего внимания, так как они похожи при беглом взгляде на "пеньки" отломавшихся побегов, и поэтому в расчет нами не принимались. Однако именно с этой категорией аномалий связаны самые настоящие разнообразные уродства, которые здесь мы только условно обозначаем в виде сложной аномалии 111—1, описанной ниже. Кроме нее, среди ювенильных уродцев часто встречаются еще три других, описанных выше, а именно 11—1, 11-11, 11-30 и одна описанная ниже — VII—1.
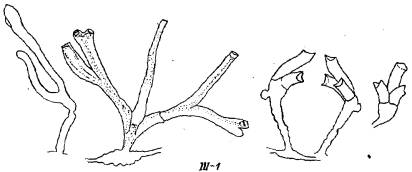
Рис. 5. Ювенильные уродливые побеги Dynamena pumila (группа III)
111—1. Уродливое развитие всего междоузлия побега. Содержит целую совокупность взаимосвязанных отклонений от нормы в развитии различных элементов первого междоузлия побега. Верхушка расчленяется на 2, реже 3 зачатка. В данном случае левая на рисунке гидротека не только необычной формы, но и имеет латеральный отросток, что совершенно не свойственно нормальному морфогенезу гидрантов. Правый зачаток дает необычную ветвь с удлиненными гидротеками, расширяющимися к устью, причем без диафрагм и с нетипичным для D. pumila изгибом в сагиттальной (перпендикулярно.й фронтальной) плоскости. О частоте встречаемости подобных аномалий пока судить преждевременно.
Группа IV: Отклонения от нормы строения гидротек (рис. 6)
После подразделения в процессе нормального роста верхушки побега на 3 зачатка (см. рис. 2) дальнейшее развитие гидрантов в значительной степени протекает самостоятельно. Это обстоятельство позволяет рассматривать группу отклонений от нормы строения гидротек отдельно, как проявление особенностей локального морфогенеза, в основном ограниченного одним зачатком.
1V—\ (9). Гидротека на гидротеке. Аномалия представляет собой результат повторного развития гидранта над устьем имеющейся гидротеки. Такое возможно благодаря периодическому рассасыванию гидрантов и повторному их образованию. В норме этот процесс должен протекать у D. pumila внутри прежней гидротеки. Вероятно, иногда новый зачаток гидранта выходит за пределы старой гидротеки. в результате чего над ней надстраивается (в виде продолжения) вторичная гидротека. Встречается редко.
IV—2. Устье гидротеки надстроено. Гидротека почти во всех отношениях нормальной формы, с хорошо заметной границей первоначального устья. Вероятно, при повторном образовании гидранта внутри старой гидротеки устье оказалось надстроенным. Встречается редко.
IV—3. Удлиненная гидротека с нормально расположенной диафрагмой. Гидротека удлиненной формы по сравнению с парной, расположенной супротивно к ней. Диафрагмы обеих гидротек расположены на одном уровне. Это означает, что аномалия не связана с этапом расчленения верхушки на 3 зачатка, а проявляется на более позднем этапе морфогенеза. Встречается периодически.
IV—4. Удлиненная гидротека со смещенным положением диафрагмы. Удлиненная гидротека в целом очень похожа на аномалию IV—3, за тем исключением, что ее диафрагма образуется несколько выше нормального уровня. По положению нижнего края внутренней стенки гидротеки можно заключить, что расчленение верхушки роста на 3 зачатка проходит нормально, хотя один из крайних зачатков остается дольше соединенным прилежащей сторой со средним, что. возможно, и приводит к подобной аномалии. Встречается периодически.
IV—5. Удлиненная гидротека оказывается частью последующего междоузлия. Аномалия представляет собой предельно выраженный вариант предыдущей
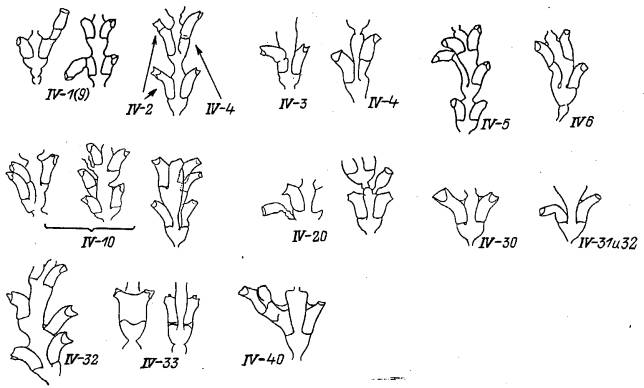
Рис. 6. Отклонения от нормы строения гидротек Dynamena pumila (группа IV)
аномалии. Принципиальная особенность заключается в том, что один из зачатков гидрантов, связанный со стволом побега дольше обычного, оказывает влияние на положение верхушки роста. Встречается редко.
IV—6. Удлиненная гидротека замещает аналогичную в последующем междоузлии. Зачаток гидранта, не отделившийся в процессе роста верхушки побега от ствола в полной мере, остается частью верхушки роста, непосредственно влияя на дальнейшее расщепление ее на зачатки. Встречается очень редко.
Все варианты “удлиненных гидротек” вместе составляют значительную долю в общей совокупности аномалий, встречаясь часто.
IV—10. Дополнительная гидротека в плоскости междоузлия. В пазухе гидротеки развивается еще одна гидротека. Это происходит, очевидно, в результате неоднократного расщепления верхушки роста на зачатки, что бывает в двух случаях: 1) если верхушка слишком крупная; 2) если деление на зачатки происходит не одномоментно: сначала отделяется бороздой один зачаток, а позже оставшаяся большая часть верхушки претерпевает повторное расщепление уже на 3 зачатка. Встречается редко.
IV—20. Гидротека расположена в необычном месте и обладает ножкой. Гидрант образуется необычным для D. pumila путем — от почки, а не в результате продольного деления верхушки роста. В результате форма гидротеки несколько отличается от нормальной. Почкованием на побеге у D. pumila в норме образуются гонангии и боковые ветви. Оказывается, не исключено образование таким же образом и гидрантов, что можно рассматривать как атавизм. Встречается редко.
IV—30. Гидротека, сохраняя нормальное положение в междоузлии, уродливой формы. Гидротека крупнее нормальной и с расширенной к устью дисталь-ной частью. Встречается очень редко.
IV—31. Дистальная часть гидротеки изогнута вниз. Гидротека в основном нормальной формы, однако ее дистальная часть по каким-то причинам отклонена вниз. Встречается редко.
IV—32. Прилежащая к стволу побега сторона гидротеки свободна почти на всем ее протяжении. Очевидно, после расчленения верхушки на 3 зачатка один из боковых рано потерял связь с центральным. Образовавшаяся гидротека торчит вбок. Вдобавок, дистальная часть гидротеки изогнута вниз. Встречается редко.
IV—33. Гидротека имеет два ус т ь я. У основания гидротека мало отличается от нормы, но к устью она необычно расширяется, раздваивается и имеет два более или менее нормальных устья. В том случае, если данная аномалия встречается в ряду последовательной трансформации междоузлии от однорядного расположения гидрантов к двурядному или же в обратной последовательности, то тогда она рассматривается как часть комплексной аномалии № 24 или №25.
IV—40. Развитие гидротеки завершается образованием междоузлия однорядного побега. Зачаток гидранта, очевидно, был большего, чем в норме, размера, и развитие гидротеки завершилось расщеплением зачатка на два и формированием междоузлия однорядного побега. В отличие от аномалии V—50(11) формирование побега гидротеки произошло исходно, а не вторично. В отличие от аномалии V—40 зачаток гидранта стал развиваться в боковую ветвь не сразу. Встречается единично.
Группа V: Отклонения от нормы расположения боковых ветвей побега (рис. 7)
Боковые побеги образуются у D. pumila в норме поодиночке сбоку междоузлия от почек ниже диафрагмы гидротеки (см. рис. I). Оси боковых ветвей должны лежать в плоскости побега. Все случаи отклонения от этого правила считаются аномалиями.
V—1(15). Боковой побег растет перпендикулярно фронтальной плоскости основного побега. Боковой побег образуется в плоскости, перпендикулярной фронтальной. Растет он от нижней части междоузлия. По форме нормален. Встречается часто.
V—10(16). Два боковых побега рядом. Оба побега нормальны по форме и растут из положенного места. Аномально то, что из одного места образуются 2 побега. В норме 2 побега из 1 междоузлия могут образовываться только супротивно. Встречается периодически.
V—20. Боковой побег растет из стенки гидротеки. Образование боковой почки на гидранте маловероятно. Скорее, почка возникла при повторном росте зачатка гидранта после рассасывания предыдущего. На это косвенно указывает сочетание данной аномалии с надстроенной гидротекой (аномалия IV—2). Встречается очень редко.
V—30 (7). Боковая ветвь растет из пазухи гндротеки. Очевидно, данная аномалия возникла в результате преждевременного расчленения верхушки побега на 3 зачатка. каждый из которых сохранил морфогенетические свойства верхушки побега. Так образовались три основных ствола, хотя два боковых по своему положению должны считаться ветвями побега. Встречается очень редко.
V—40. Боковая ветвь вместо гидротеки. Зачаток гидранта по каким-то причинам сохранил морфогенетические свойства верхушки побега. С самого начала на месте гидранта стал формироваться побег. На это указывают отделенность всего образования от ствола основного побега, отсутствие каких-либо морфогенетических признаков строения гидротеки, в том числе отсутствие диафрагмы. Встречается редко.
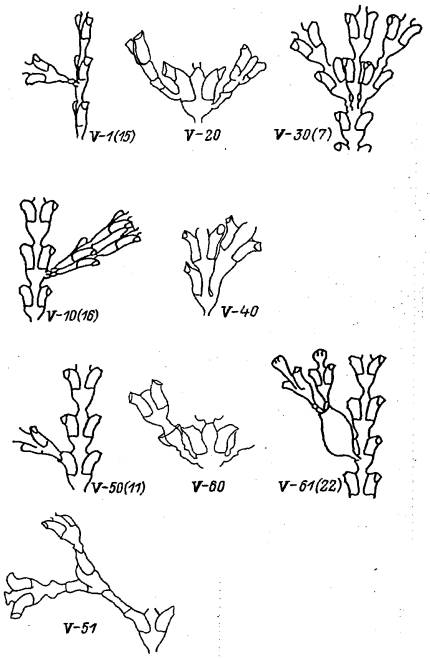
Рис. 7. Отклонения от нормы расположения боковых ветвей побега Dynamena pumila (группа V)
V—50(11). Боковая ветвь растет из устья гидротеки. На гидротеке, из которой выросла боковая ветвь, видны ясные следы устья. Это означает, что боковая ветвь возникла после рассасывания гидранта. При повторном росте зачатка гидранта образовалась ветвь. Встречается часто.
V—51. Усложненный вариант боковых ветвей из гидротек. Боковые ветви образуются из гидротек последовательно неоднократно. Встречается редко.
V—60. Боковая ветвь побега растет из гонотеки. Из недосформировавшейся гонотеки выросла боковая ветвь побега. Хорошо видно, что ее образование произошло после того, как гонангий рассосался. Внутри гонотеки и вне связи с ней при повторном росте зачатка образовалась боковая ветвь. Встречается редко.
V—61 (22). Побег от верхушки полноценной гонотеки. На дистальной части пустой гонотеки выросли два молодых побега. Побеги, по всей вероятности, образовались в результате прорастания планул. Побеги связаны с материнским побегом гидроризами, которые проходят внутри гонотеки. Этот вариант роста побегов из гонотеки нельзя рассматривать как аномалию. Это хороший пример артефакта. Встречается очень редко.
Группа VI: Отклонения от нормы формы и расположения гонотек (рис. 8)
В нашем материале было мало колоний с гонотеками. Обычно гонангий развиваются только в короткий период размножения, а затем пустые гонотеки отваливаются. В период размножения колонии растут плохо, поэтому мы не собирали материал в это время.
VI—1(19). Боковой побег в виде триады гонотек. В боковом побеге после расчленения верхушки на 3 зачатка из каждого зачатка образовалась гонотека. Из 3 гонотек центральная более крупная.
Рис.8. Отклонения от нормы формы и расположения гонотек Dynamena pumila (группа IV)
 |
Все гонотеки не отличаются от нормальных,находящихся на той же стадии развития. Аномалия возникла в период интенсивного образования гонотек на колонии, в которой этот процесс протекал особенно бурно. Аномалия была обнаружена лишь единично ранее (Марфенин, 1975) и не была найдена в исследованной теперь обширной выборке.
Группа VII: Отклонения от нормы положения столонов в колонии (рис. 9)
По опыту экспериментальной работы нам известно, что перерождение любых зачатков (верхушки роста побега, боковой ветви, гидранта) в столон — обычное явление для Dynamena pumila в том случае, если ее побеги по какой-то причине прижаты к субстрату. поэтому подобные морфогенетические реакции можно считать свойственными виду, а соответствующие аномалии — морфотипами вида.
VII—1 (17). Верхушка побега перерождается в столон. Столон, как правило, растет, плавно изгибаясь в одну сторону, что увеличивает вероятность его контакта с субстратом. Встречается редко.
VII—10. Столон растет от ствола побега. Чаще всего возникает на месте боковой ветви, т. е. от почки. Встречается редко.
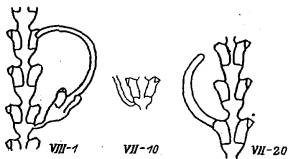
Рис. 9. Отклонения от нормы положения столонов в колонии Dynamena pumila (группа VII)
VII—20( 10). Столон растет из гидротеки. На гидротеке видны ясные следы устья. Это означает, что в гидротеке был нормальный гидрант, который рассосался. При повторном росте зачатка образовался столон. Встречается периодически.
ПРОБЛЕМА СИСТЕМАТИЗАЦИИ АНОМАЛИЙ И МОРФОТИПОВ
Предложенная выше простейшая классификация аномалий. естественно, не является единственно приемлемой или даже лучшей. Всякая систематизация обретает свой смысл только относительно рассматриваемой задачи. В данном случае предполагается использовать в дальнейшем фототипические вариации строения побега Dynamena pumila для трех целей: 1) количественного анализа популяционной изменчивости вида; 2) изучения реакции вида на различные факторы среды; 3) поисков пределов морфогенетических возможностей вида.
Первые две задачи допускают достаточно формальную систематизацию аномалий, лишь бы классификация была не громоздкой. а кластеры были достаточно ясно разграничены между собой, чтобы легко можно было применять количественные методы сравнения, поэтому излишняя детализация для достижения данной цели противопоказана.
Для третьей задачи — анализа широты морфогенетических возможностей вида, напротив, целесообразнее скрупулезно выделить в спектре морфотипов все множество вариантов строения, а классификацию проводить в соответствии и с учетом предполагаемых механизмов морфогенеза. В этом случае количество описанных выше аномалий D. pumila должно быть увеличено примерно вдвое.
Мы выбрали первый вариант классификации, постарались ограничить разнообразие аномалий, ориентируясь на определяющие признаки, указанные в описании аномалий, поэтому при практическом использовании нашей классификации рекомендуется множество принципиально не различающихся разновидностей сводить к ключевому морфотипу, искусственно занижая, таким образом, морфогенетическую вариабельность.
Подразделение всей совокупности аномалий на семь разновеликих групп, выделенных достаточно условно, позволяет при необходимости, располагать в системе новые нахддки ближе к родственным аномалиям. Той же цели служит дробная нумерация (1...10...20...), в которой зарезервированы обозначения для еще не обнаруженных морфотипов.
Нумерация предшествующей классификации (Марфенин, 1975) также сохранена в виде числа в скобках. Это не только служит обеспечению преемственности: оказалось, что все наиболее часто встречаемые аномалии были представлены в прежней компактной системе. Редко встречаемые аномалии могут быть для решения многих задач сведены вместе или оставлены без рассмотрения как не достоверные для количественного сравнения, тогда удобнее пользоваться более простой старой классификацией, однако для обработки нового фактического материала правильнее применять предлагаемую усовершенствованную и существенно пополненную классификацию. Все предложения по дальнейшему совершенствованию системы морфотипов Dynamena риmila просим присылать по адресу: 119899, Москва, Московский государственный университет, биологический факультет, кафедра зоологии беспозвоночных, тел.: 939-36-56.
Список литературы
Марфенин Н. Н. Аномалии формы побега колонки Dynamena pumila (Hydrozoa, Leptolida). // Комплексные исследования природы океана, вып. 5.—М., 1975. С. 230—239.
Марфенин Н. Н. Морфологические аномалии — резерв адаптивной радиации (на примере колониальных гидроидов) // Следы жизнедеятельности и динамика среды в древних биотопах: Тр. XXX сессии Всесюзн. палеонтол. о-ва.— Киев, 1988. С. 149—156.