| Скачать .docx | Скачать .pdf |
Реферат: Нуклеиновые кислоты
Виды нуклеиновых кислот. Нуклеиновые кислоты — фосфорсодержащие биополимеры живых организмов, обеспечивающие хранение и передачу наследственной информации. Они были открыты в 1869 г. швейцарским биохимиком Ф. Мишером в ядрах лейкоцитов, сперматозоидов лосося. Впоследствии нуклеиновые кислоты обнаружили во всех растительных и животных клетках, вирусах, бактериях и грибах.
В природе существует два вида нуклеиновых кислот — дезок-сирибонуклеиновые (ДНК) и рибонуклеиновые (РНК). Различие в названиях объясняется тем, что молекула ДНК содержит пяти-углеродный сахар дезоксирибозу, а молекула РНК— рибозу. В настоящее время известно большое число разновидностей ДНК и РНК, отличающихся друг от друга по строению и значению в метаболизме.
ДНК находится преимущественно в хромосомах клеточного ядра (99% всей ДНК клетки), а также в митохондриях и хлоропластах. РНК входит в состав рибосом; молекулы РНК содержатся также в цитоплазме, матриксе пластид и митохондрий.
Нуклеотиды — структурные компоненты нуклеиновых кислот. Нуклеиновые кислоты представляют собой биополимеры, мономерами которых являются нуклеотиды.
Нуклеотиды —сложные вещества. В состав каждого нуклео-тида входит азотистое основание, пятиуглеродный сахар (рибоза или дезоксирибоза) и остаток фосфорной кислоты.
Существует пять основных азотистых оснований: аденин, гуанин, урацил, тимин и цитозин. Первые два являются пуриновыми; их молекулы состоят из двух колец, первое содержит пять членов, второе — шесть. Следующие три являются пиримидинами и имеют одно пятичленное кольцо.
Вот как выглядит, например, формула тимидилового нуклеотида(тимидин):
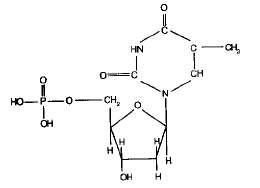
Названия нуклеотидов происходят от названия соответствующих азотистых оснований; и те и другие обозначаются заглавными буквами: аденин — аденилат (А), гуанин — гуанилат (Г), цитозин — цитидилат (Ц), тимин — тимидилат (Т), урацил — уридилат (У).
Количество нуклеотидов в молекуле нуклеиновых кислот бывает разным — от 80 в молекулах транспортных РНК до нескольких сотен миллионов у ДНК.
ДНК. Молекула ДНК состоит из двух полинуклеотидных, спирально закрученных относительно друг друга цепочек.
В состав нуклеотидов молекулы ДНК входят четыре вида азотистых оснований: аденин, гуанин, тимин и цитоцин. В полинук-леотидной цепочке соседние нуклеотиды связаны между собой ковалентными связями, которые образуются между фосфатной группой одного нуклеотида и З'-гидроксильной группой пентозы другого. Такие связи называются фосфодиэфирными. Фосфатная группа образует мостик между З'-углеродом одного пентоз-ного цикла и 5-углеродом следующего. Остов цепей ДНК образован, таким образом, сахарофосфатными остатками (рис. 1.2).
Хотя в состав ДНК входит четыре типа нуклеотидов, благодаря различной последовательности их расположения в длинной цепочке достигается огромное разнообразие этих молекул.
Полинуклеотидная цепь ДНК закручена в виде спирали наподобие винтовой лестницы и соединена с другой, комплементарной ей цепью с помощью водородных связей, образующихся между адени-ном и тимином (две связи), а также гуанином и цитозином (три связи). Нуклеотиды А и Т, Г и Ц называются комплементарными.
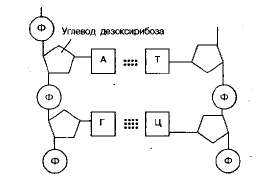
Рис 1.2. Фрагмент молекулы ДНК (между А—Т— две водородные связи; между Г—Ц — три водородные связи).
В результате у всякого организма число адениловых нуклеотидов равно числу тимидиловых, а число гуаниловых — числу цитидиловых. Эта закономерность получила название «правило Чаргаффа». Благодаря этому свойству последовательность нуклеотидов в одной цепи определяет их последовательность в другой. Такая способность к избирательному соединению нуклеотидов называется комплемен-тарностью, и это свойство лежит в основе образования новых молекул ДНК на базе исходной молекулы (репликации, т. е. удвоения).
Цепи в молекуле ДНК противоположно направлены (антипа-раллелъностъ). Так, если для одной цепи мы выбираем направление от З'-конца к 5'-концу, то вторая цепь с таким направлением будет ориентирована противоположно первой — от 5-конца к З'-концу, иначе говоря, «голова» одной цепи соединяется с «хвостом» другой и наоборот.
Впервые модель молекулы ДНК была предложена в 1953 г. американским ученым Дж. Уотсоном и англичанином Ф. Криком на основе данных Э. Чаргаффа о соотношении пуриновых и пиримидиновых оснований молекул ДНК и результатов рентге-но-структурного анализа, полученных М. Уилкинсом и Р. Франклин. За разработку двухспиральной модели молекулы ДНК Уот-сон, Крик и Уилкинс были удостоены в 1962 г. Нобелевской премии.
ДНК — самые крупные биологические молекулы. Их длина составляет от 0,25 (у некоторых бактерий) до 40 мм (у человека). Это значительно больше самой крупной молекулы белка, которая в развернутом виде достигает длины не более 100—200 нм. Масса молекулы ДНК составляет 6x10-12 г.
Диаметр молекулы ДНК 2 нм, шаг спирали 3,4 нм; каждый виток спирали содержит 10 пар нуклеотидов. Спиральная структура поддерживается многочисленными водородными связями, возникающими между комплементарными азотистыми основаниями, и гидрофобными взаимодействиями. Молекулы ДНК эука-риотических организмов линейны. У прокариот ДНК, напротив, замкнута в кольцо и не имеет ни 3-, ни 5-концов.
При изменении условий ДНК, подобно белкам, может под-. вергаться денатурации, которая называется плавлением. При постепенном возврате к нормальным условиям ДНК ренатурирует.
Функцией ДНК является хранение, передача и воспроизведение в ряду поколений генетической информации. В ДНК любой клетки закодирована информация обо всех белках данного организма, о том, какие белки, в какой последовательности и в каком количестве будут синтезироваться. Последовательность аминокислот в белках записана в ДНК так называемым генетическим (триплетным) кодом.
Основным свойством ДНК является ее способность к репликации.
Репликация — это процесс самоудвоения молекул ДНК, происходящий под контролем ферментов. Репликация осуществляется перед каждым делением ядра. Начинается она с того, что спираль ДНК временно раскручивается под действием фермента ДНК-полимеразы. На каждой из цепей, образовавшихся после разрыва водородных связей, по принципу комплементарности синтезируется дочерняя цепь ДНК. Материалом для синтеза служат свободные нуклеотиды, которые есть в ядре (рис. 1.3).
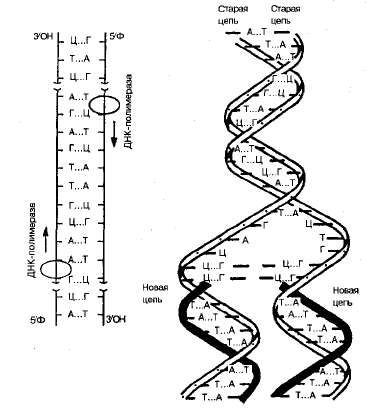
Рис. 1.3.. Схема репликации ДНК
Таким образом, каждая полинуклеотидная цепь выполняет роль матрицы для новой комплементарной цепи (поэтому процесс удвоения молекул ДНК относится к реакциям матричного синтеза). В результате получается две молекулы ДНК, у каждой из которых ' одна цепь остается от родительской молекулы (половина), а другая — вновь синтезированная. Причем одна новая цепь синтезируются сплошной, а вторая — сначала в виде коротких фрагментов, которые затем сшиваются в длинную цепь специальным ферментом—ДНК-лигазой. В результате репликации две новые молекулы ДНК представляют собой точную копию исходной молекулы.
Биологический смысл репликации заключается в точной передаче наследственной информации от материнской клетки к дочерним, что и происходит при делении соматических клеток.
РНК. Строение молекул РНК во многом сходно со строением молекул ДНК. Однако имеется и ряд существенных отличий. В молекуле РНК вместо дезоксирибозы в состав нуклеотидов входит рибоза, вместо тимидилового нуклеотида (Т) — уридило-вый (У). Главное отличие от ДНК состоит в том, что молекула РНК представляет собой одну цепь. Однако ее нуклеотиды способны образовывать водородные связи между собой (например, в молекулах тРНК, рРНК), но в этом случае речь идет о внутри-цепочечном соединении комплементарных нуклеотидов. Цепочки РНК значительно короче ДНК.
В клетке существует несколько видов РНК, которые различаются по величине молекул, структуре, расположению в клетке и функциям:
Информационная (матричная) РНК(иРНК). Этот вид наиболее разнороден по размерам и структуре. иРНК представляет собой незамкнутую полинуклеотидную цепь. Она синтезируется в ядре при участии фермента РНК-полимеразы, комплементарна участку ДНК, на котором происходит ее синтез. Несмотря на относительно низкое содержание (3—5% РНК клетки), она выполняет важнейшую функцию в клетке: служит в качестве матрицы для синтеза белков, передавая информацию об их структуре с молекул ДНК. Каждь|й белок клетки кодируется специфической иРНК, поэтому число их типов в клетке соответствует числу видов белков.
Рибосомная РНК (рРНК). Это одноцепочечные нуклеиновые кислоты, образующие в комплексе с белками рибосомы — орга-неллы, на которых происходит синтез белка. Рибосомные РНК синтезируются в ядре. Информация об их структуре закодирована в участках ДНК, которые расположены в области вторичной перетяжки хромосом. Рибосомные РНК составляют 80% всей РНК клетки, поскольку в клетке имеется огромное количество рибосом. Рибосомные РНК обладают сложной вторичной и третичной структурой, образуя петли на комплементарных участках, что приводит к самоорганизации этих молекул в сложное по форме тело. В состав рибосом входит три типа рРНК у прокариот и четыре типа рРНК у эукариот. 3. Транспортная (трансферная) РНК(тРНК). Молекула тРНК состоит в среднем из 80 нуклеотидов. Содержание тРНК в клетке — около 15% всей РНК. Функция тРНК — перенос аминокислот к месту синтеза белка. Число различных типов тРНК в клетке невелико (20—60). Все они имеют сходную пространственную организацию. Благодаря внутрицепочечкым водо-родным связям молекула тРНК приобретает характерную вторичную структуру, называемую клеверным листам. Трехмерная же модель тРНК выглядит несколько иначе. В тРНК выделяют четыре петли: акцепторную (служит местом присоединения аминокислоты), антикодоновую (узнает кодон в иРНК в процессе трансляции) и две боковые.
Источник : Н.А. Лемеза Л.В.Камлюк Н.Д. Лисов "Пособие по биологии для поступающих в ВУЗы"