| Скачать .docx | Скачать .pdf |
Реферат: Механика митоза
Л.В. Яковенко
Вскоре после того, как немецкий патофизиолог Р.Вирхов в середине XIX в. сформулировал основной принцип клеточной теории в виде афоризма Omni cellula ex cellula («Всякая клетка – из другой клетки»), было установлено, что жизнь соматической клетки протекает циклически, начинаясь с деления и делением оканчиваясь. За полтора века, прошедшие с тех пор, получено множество новых данных об особенностях деления различных клеток. Стали понятны многие процессы организации и регуляции деления, их невероятная сложность. И все большее восхищение исследователей вызывает точность, с которой происходит разделение хромосом между будущими дочерними клетками. Именно о механизмах разделения хромосом (на примере клеток животных) и пойдет речь ниже.
Клеточный цикл – это последовательность закономерно сменяющих друг друга фаз от образования клетки в результате деления до либо разделения ее на дочерние клетки в следующем акте деления, либо гибели. У эукариот клеточный цикл состоит из интерфазы и собственно деления, или митоза. Каждой из этих фаз соответствуют определенные явления и процессы, которые позволяют разделить их на более мелкие стадии. У разных организмов количество и последовательности стадий клеточного цикла различаются.
Интерфаза значительно более длительна, чем митоз (обычно занимает не менее 90% всего времени клеточного цикла), и обычно подразделяется на три периода: пресинтетический (G1), синтетический (S) и постсинтетический (G2). На стадии G2 клетка может перейти к следующему делению или к состоянию покоя (G0). Переход к делению возможен только из стадии G2, поэтому, если клетка находится в состоянии G0, для продолжения деления ей необходимо вернуться в состояние G2. Стадия G1 может продолжаться от 2 ч до нескольких недель или даже месяцев, продолжительность стадии S 6–12 ч, а стадии G2 – от получаса до нескольких часов.
Собственно непрямое деление, или митоз, состоит из стадий кариокинеза (деления ядра) и цитокинеза (деления цитоплазмы). Разделение хромосом происходит на стадии кариокинеза, поэтому рассмотрим ее подробнее.
В первой фазе митоза – профазе – хромосомы спирализуются и становятся видны в световой микроскоп в виде тонких нитей. Клеточные центры, удвоение которых происходит на стадии S, расходятся к полюсам клетки. В конце профазы ядрышки исчезают, ядерная оболочка разрушается и хромосомы выходят в цитоплазму.
Затем клетка переходит в метафазу, начало которой называют прометафазой. В прометафазе хромосомы располагаются в цитоплазме довольно беспорядочно. Формируется митотический аппарат, в состав которого входит веретено деления и центриоли. Веретено деления – это система особых структур, микротрубочек (МТ), в делящейся клетке, обеспечивающая расхождение хромосом. Затем кинетохоры (центромеры) хромосом захватываются МТ, отходящими от обоих полюсов веретена деления, и через некоторое время хромосомы выстраиваются в экваториальной плоскости клетки. В метафазе хромосомы максимально спирализованы. Центромеры хромосом располагаются в экваториальной плоскости клетки независимо друг от друга. Совокупность хромосом в экваториальной плоскости клетки образует метафазную пластинку.
Наша справка Митоз (от греч. mitos — нить) – непрямое деление клетки, наиболее распространённый способ воспроизведения клеток, обеспечивающий тождественное распределение генетического материала между дочерними клетками и преемственность хромосом в ряду клеточных поколений. Биологическое значение митоза определяется сочетанием в нём удвоения хромосом путём продольного расщепления их и равномерного распределения между дочерними клетками. |
На следующей стадии деления – в анафазе – происходит разделение хромосом на хроматиды. С этого момента каждая хроматида становится самостоятельной однохроматидной хромосомой. Сначала сестринские хроматиды расходятся к противоположным полюсам веретена деления, а сами полюса остаются неподвижными (анафаза А), а затем полюса веретена расходятся к противоположным концам клетки (анафаза В).
После этого клетка переходит в телофазу: веретено деления разрушается, хромосомы у полюсов клетки деспирализуются, вокруг них формируются ядерные оболочки. В клетке образуются два ядра, генетически идентичные исходному ядру.
С окончанием кариокинеза клетка переходит в стадию цитокинеза, на которой происходит разделение цитоплазмы и формирование мембран дочерних клеток. У животных цитокинез происходит путем «перешнуровывания» клетки. У растений цитокинез происходит иначе: в экваториальной плоскости образуются пузырьки, которые сливаются с образованием двух параллельных мембран. На этом митоз завершается, и дочерние клетки переходят в интерфазу.
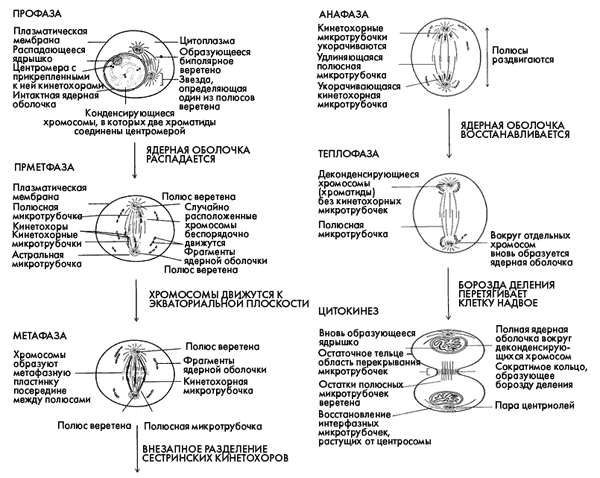
Шесть стадий клеточного деления
На всех стадиях кариокинеза важнейшую роль играют МТ – их образование и пространственная ориентация, взаимодействие с кинетохорами хромосом, структурные изменения, создающие силы, необходимые для разделения хромосом, и, наконец, их разрушение. МТ входят в состав цитоскелета и играют важнейшую роль в поддержании и изменении формы клетки и направленном переносе внутриклеточных компонентов (везикул, органелл, белков и т.п.) в цитоплазме. В клетках животных несколько тысяч МТ. Все они растут из специальных образований, называемых центрами организации МТ (ЦОМТ). В клетке может быть 1–2 ЦОМТ. Исследования показали, что от центросомы отходят всего несколько десятков МТ, следовательно, МТ не обязательно связаны с центросомой. Центриоли же дают начало новым МТ, которые приходят на смену постепенно деполимеризующимся старым.
Центросома, или клеточный центр, – главный ЦОМТ и регулятор хода клеточного цикла в клетках эукариот. Центросома состоит из аморфного материала и пары центриолей – материнской и дочерней, расположенных строго определенным образом и образующих структуру, называемую диплосомой. (О структуре и функциях центросом можно прочитать, например, в журнале «Природа», 2007, №5.) Помимо участия в делении ядра, центросома играет важную роль в формировании жгутиков и ресничек. Центриоли, расположенные в ней, выполняют функцию центров организации для МТ аксонем жгутиков. У организмов, лишенных центриолей (например, у сумчатых и базидиевых грибов, покрытосеменных растений), жгутики не развиваются.
ЦОМТ могут репродуцироваться самостоятельно: новый центр образуется рядом с существующим, а затем отходит от него. До сих пор оставалось тайной, как это происходит. Но совсем недавно американские ученые, изучая экстракты центросом ооцитов моллюска Spisula solidissima, обнаружили, что центросомы содержат особые молекулы РНК. Учитывая, что центросомы имеют очень древнее происхождение и чрезвычайно консервативны, это открытие позволило предположить, что они имеют собственный генетический аппарат.
Наша справка Продолжительность митоза зависит от размеров клеток, их плоидности, числа ядер, а также от условий окружающей среды, в частности от температуры. В животных клетках митоз длится 30–60 мин, в растительных – 2–3 ч. Большинству клеток присущ суточный ритм деления. В органах ночных животных максимум митотических делений отмечается, как правило, утром, а минимум — в ночное время. У дневных животных и человека отмечается обратная динамика суточного ритма. |
МТ представляет собой очень маленькую трубочку длиной несколько микрометров при наружном диаметре 25 нм. Она построена из 13 длинных «палочек» – протофиламентов, параллельных оси трубочки и расположенных по кругу. Протофиламент составлен из чередующихся глобул альфа- и бета-тубулина, причем в каждой паре таких глобул (димере тубулина) альфа-тубулин взаимодействует с бета-тубулином, а бета-тубулин – с альфа-тубулином ближайших соседних димеров, что и позволяет образоваться очень прочной цилиндрической конструкции. Как же такая конструкция может обеспечивать перемещение чего-либо внутри клетки?
Что касается органелл, белков и других компонентов клетки, то они перемещаются по МТ, прикрепляясь к белкам-моторам: динеинам и кинезинам, которые способны буквально «шагать» по МТ в определенном направлении, потребляя в качестве топлива АТФ. Хромосомы же прикрепляются к концам МТ, которые затем каким-то образом быстро растаскивают их к полюсам веретена деления.
Было известно, что длина МТ может быть постоянной, как, например, в жгутиках. Однако длина цитоплазматических МТ меняется постоянно: они то растут, то укорачиваются, могут исчезнуть совсем, потом опять начнут расти… Когда МТ в процессе роста достигает мишени, ее длина стабилизируется, но как это происходит, до сих пор не вполне ясно.
Экспериментально установлено, что МТ может находиться в трех основных состояниях: полимеризации, деполимеризации и катастрофы. Полимеризация – это присоединение одиночных молекул тубулина, находящихся в цитоплазме, к торцу трубочки (деполимеризация – обратный процесс). Альфа- и бета-субъединицы димера тубулина в цитоплазме сначала присоединяют по одной молекуле гуанозинтрифосфата (ГТФ), похожего по свойствам на АТФ, а затем уже могут присоединиться к торцу растущей МТ. Для роста МТ необходимо также наличие в цитоплазме некоторых специфических белков, присутствие ионов магния и отсутствие ионов кальция.
Пока с димером тубулина связаны две молекулы ГТФ, он находится в Т-состоянии, и при этом вся конструкция трубочки устойчива. Однако на бета-субъединице димера тубулина через некоторое время происходит гидролиз ГТФ, который превращается в гуанозиндифосфат (ГДФ), при этом весь димер переходит в D-состояние, а кольцо молекул тубулина на торце МТ становится напряженным, неустойчивым. В этом состоянии к торцу МТ уже не могут присоединиться новые димеры тубулина, и МТ переходит в состояние катастрофы. Поэтому рост МТ возможен только пока на конце МТ есть кольцо из Т-димеров тубулина, так называемая Т-шапочка. Если концентрация тубулина в цитоплазме невелика, димеры «Т-шапочки» могут успеть перейти в D-состояние, прежде чем к ним присоединятся новые Т-димеры и трубочка перейдет в состояние катастрофы.
Если при деполимеризации происходит отсоединение молекул тубулина по кольцу на торце МТ, то при катастрофе протофиламенты разъединяются, как отдельные проволочки, и стремятся закрутиться в колечки. При этом разборка МТ происходит очень быстро. Конец МТ, закрепленный в центросоме и защищенный от катастроф, называют «минус»-концом МТ, а другой конец, который либо нарастает, либо быстро разрушается – «плюс»-концом. В цитоплазме существует множество белков, которые могут взаимодействовать с тубулином в разных состояниях, влияя на скорость роста или распада МТ. Существенно, что белки-моторы умеют различать «плюс»- и «минус»-концы МТ: динеины движутся к «минус»-концу, а кинезины – к «плюс»-концу микротрубочки.
Каждой стадии митоза соответствует особое поведение МТ. Митотическое деление происходит с образованием специальной структуры – веретена деления, основой строения которого являются МТ, исходящие из двух клеточных центров, расположенных в полюсах клетки. Веретено деления состоит как бы из двух перекрывающихся в центральной части полуверетен, на концах которых находятся центросомы. В растительных клетках образование веретена деления происходит без участия центросом.Всего можно выделить три типа МТ: астральные, полюсные и кинетохорные. Кинетохорные МТ связывают центросому с кинетохором хромосомы. Они образуются в прометафазе. На стадии ранней профазы быстро растут астральные МТ, направленные радиально от каждого из двух клеточных центров. Астральные МТ тянутся от центросом к периферии клетки, их «плюс»-концы взаимодействуют с белками, закрепленными в клеточной мембране, по-видимому, с помощью динеинов, притягивающих центросомы к мембране.
В это же время появляются полюсные МТ, которые растут по направлению от одного клеточного центра к другому. Полюсные МТ имеют тенденцию объединяться в группы от двух до шести МТ (на стадии метафазы), в основном с МТ противоположного полюса. Так образуются полюсные нити, в которых МТ направлены антипараллельно, т.е. «плюс»-концами в противоположные стороны. Упомянутые выше моторные белки, взаимодействуя с антипараллельными МТ, приводят либо к стягиванию клеточных центров по направлению друг к другу или к их расталкиванию. Отсутствие или дефекты какого-либо из этих моторных белков приводят к нарушениям расхождения центросом и митоза в целом.
Кроме изменений в организации МТ, связанной с удвоением центросомы, изменяется и их динамика. Во время интерфазы МТ относительно длинные и стабильные, состояние роста длится в среднем около 10 мин. При переходе к митозу частота катастроф увеличивается примерно в 10 раз, поэтому состояние роста МТ укорачивается и становится меньше 1 мин. Эти изменения вызываются, в основном, специальными белками, контролирующими ход митоза, и приводят к тому, что МТ становятся нестабильными, быстро изменяющимися.
Благодаря тому, что на стадии прометафазы ядерная мембрана уже разрушена, МТ могут дотянуться до хромосом. Присоединение их к кинетохорам происходит случайно, при соприкосновении кинетохора с «плюс»-концом или боковой поверхностью МТ. В последнем случае (латеральное взаимодействие) хромосома начинает быстро, со скоростью 20–25 мкм/мин, двигаться к соответствующему полюсу веретена деления. Эта скорость сравнима со скоростью перемещения динеина вдоль МТ, но прямых данных об участии динеина в этом процессе пока нет. Затем латеральное взаимодействие заменяется концевым за счет разрушения МТ в кинетохоре, и длина МТ стабилизируется.
Кинетохор представляет собой трехслойную структуру, видимую на микрофотографиях как два темных слоя, разделенных светлым промежутком. Он имеет длину 0, 3–0, 6 мкм и толщину около 0, 1 мкм. Один темный слой кинетохора связан с центромерой, другой – с МТ. К кинетохору могут быть прикреплены и МТ, не связанные с центросомой (в растительных и некоторых других клетках веретено деления образуется вообще без центросом). Полярность присоединения таких МТ та же: «плюс»-конец присоединен к кинетохору, а «минус»-конец находится вблизи полюса веретена. Такие МТ более стабильны, чем МТ, заканчивающиеся в полюсах веретена деления.
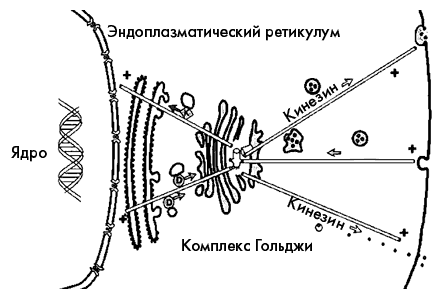
Направленный транспорт белков внутри клетки
В начале митоза кинетохоры хромосом расположены несимметрично относительно полюсов веретена деления, поэтому они быстрее захватываются МТ, идущими из ближайшего полюса. Однако до тех пор, пока сестринский кинетохор не будет захвачен МТ, идущей от другого полюса, и пара хромосом не будет расположена по экватору веретена деления, митоз не перейдет к следующей стадии – анафазе. Это обеспечивают специальные белки, входящие в состав системы контрольных точек митоза. Таких контрольных точек в клеточном цикле несколько. Только если предыдущая стадия митоза завершена нормально, они вырабатывают сигнал готовности к продолжению митоза.
К каждому их двух кинетохоров сестринских хроматид прикрепляется по 10–40 МТ, образующих кинетохорную нить. При этом скорость присоединения МТ к кинетохорам возрастает к концу метафазы примерно в 10 раз по сравнению с прометафазой. Это объясняется тем, что уже присоединившиеся к кинетохору МТ облегчают присоединение следующих МТ. Такой процесс называется кооперативным.
Наша справка Нарушения митоза. При различных патологических процессах нормальное течение митоза нарушается. Выделяют 3 основных вида патологии: 1) повреждения хромосом (набухание, склеивание, фрагментация, образование мостов, повреждения центромеров, отставание отдельных хромосом при движении, нарушение их спирализации и деспирализации, раннее разъединение хроматид, образование микроядер; 2) повреждения митотического аппарата (задержка митоза в метафазе, многополюсный, моноцентрический и асимметричный митоз, трёхгрупповая и полая метафазы); 3) нарушения цитотомии. Патологические митозы возникают после воздействия митотических ядов, токсинов, экстремальных факторов (ионизирующее излучение, аноксия, гипотермия), при вирусной инфекции и в опухоли. Резкое увеличение числа патологических митозов типично для злокачественных опухолей. |
Основная функция веретена деления – это обеспечение правильного разделения сестринских хроматид. Для направленного движения таких больших структур, как хроматиды, необходимо действие на них значительных сил. Эксперименты показывают, что существуют несколько типов таких сил.
Сила первого типа возникает за счет непрерывного наращивания «плюс»-конца МТ и деполимеризации «минус»-конца. Эти процессы (при равенстве их скоростей) приводят к тому, что димеры тубулина непрерывно перемещаются в сторону «минус»-конца, а длина трубочки при этом не меняется. Если заблокировать присоединение тубулина на «плюс»-конце МТ (добавлением таксола), то разборка МТ в центросомах все равно продолжается и центросомы начинают двигаться по направлению к хромосомам со скоростью, определяемой скоростью деполимеризации МТ. Определение скорости перемещения тубулина по таким МТ показало, что возникающая при этом сила обеспечивает до 25% скорости движения хромосом к полюсу веретена деления в анафазе. В изолированном из яйца лягушки митотическом веретене движение хромосом полностью обеспечивается этой силой.
Силы второго типа («полярный ветер») действуют на участки хроматид, не связанные с кинетохором. Экспериментально показано, что после отрезания плеч хромосом от центромеры они начинают двигаться к экватору веретена деления со скоростью около 2 мкм/мин и в конце концов занимают положение между полюсами веретена деления. Скорее всего, эти силы обусловлены взаимодействием связанных с хроматином белков-моторов (типа кинезина) с МТ.
Наконец, сила третьего типа – это сила, с которой кинетохорная нить тянет хромосому к полюсу веретена деления. Это главная сила, обеспечивающая расхождение хромосом в анафазе. Она имеет, по-видимому, несколько составляющих. Во-первых, в состав кинетохора входят моторные белки (динеин), которые могут взаимодействовать с боковой поверхностью МТ и вызывать перемещение кинетохора в сторону центросомы. Во-вторых, в кинетохоре имеются белки, которые способны существенно влиять на скорость роста или разрушения МТ в зависимости от сигналов системы контрольной точки, белки которой также находятся в кинетохоре. После прохождения контрольной точки и перехода клетки в анафазу скорость деполимеризации МТ в кинетохоре резко возрастает. В результате МТ начинает быстро сокращаться, развивая необходимую для движения хромосомы к полюсу силу. Кроме того, натяжение кинетохорных нитей возрастает даже при постоянной их длине за счет расхождения антипараллельных участков полюсных МТ и, как результат, увеличения их длины. Сила, генерируемая за счет этого процесса, тем меньше, чем больше длина полюсных МТ: упругость МТ конечна, поэтому при увеличении длины они начинают изгибаться, и сила, раздвигающая полюсы веретена деления, уменьшается. Следовательно, чем дальше друг от друга находятся полюсы веретена деления, тем меньше расталкивающая их сила.
Баланс перечисленных выше сил приводит сначала к выстраиванию хромосом по экватору веретена деления, а затем, как следствие изменения баланса, к их расхождению к полюсам. Надо отметить, что баланс этот динамический, а не статический, поэтому даже при стабильном расположении хромосом в плоскости экватора веретена деления, они постоянно смещаются то к одному полюсу, то к другому. Скорость таких колебательных движений – 2–3 мкм/мин. Пока точной модели этих колебаний нет.
Кратко суммируем сказанное выше. Важнейшей задачей митоза является правильное разделение сестринских хромосом, которое осуществляется с помощью веретена деления. Веретено деления образуется МТ, с которыми взаимодействуют белки-моторы (динеины и кинезины), кинетохоры, центриоли, мембранные белки. Белки-моторы могут связываться с белками различных внутриклеточных структур (например, с хроматином) и обеспечивают их перемещение по МТ в одну или другую сторону, осуществляемое за счет энергии гидролиза АТФ. Перемещение хромосом обеспечивается как за счет взаимодействия МТ с белками-моторами, так и за счет процессов роста или распада МТ. При этом именно соотношение скоростей последних двух процессов, регулируемое белками системы контрольных точек, обеспечивает, в основном, и выстраивание хромосом в экваториальной плоскости, и расхождение их к полюсам веретена деления.
Хотя непосредственно измерить силы, действующие со стороны МТ на хромосомы, не представляется возможным, многие детали молекулярных механизмов этих процессов позволят выяснить их адекватные модели. В последнее время стали появляться модели, связывающие биохимические и механические процессы в ходе митоза, но решающее слово, как всегда, остается за экспериментальными исследованиями, которые еще предстоит выполнить.