| Скачать .docx | Скачать .pdf |
Реферат: Незаменимые полиненасыщенные жирные кислоты и их пищевые источники для человека
Незаменимые полиненасыщенные жирные кислоты и их пищевые источники для человека
М.И. Гладышев
Институт биофизики СО РАН, Россия
Рассматриваются структура и строение молекул жирных кислот, включая незаменимые полиненасыщенные жирные кислоты (ПНЖК). Описывается роль ПНЖК в организме человека как биохимических предшественников различных эндогормонов. Приводится обзор результатов многолетних клинических и эпидемиологических исследований действия ПНЖК на человека. Рассматривается значение сбалансированного потребления ПНЖК в диете, способствующего профилактике сердечно-сосудистых заболеваний. Обобщаются данные о содержании ПНЖК в рыбе как в основном источнике этих веществ в питании человека. Обсуждается вероятная роль потребления морепродуктов в эволюции человека. Даётся характеристика водных экосистем как основных продуцентов длинноцепочечных ПНЖК в биосфере. Представлены основные способы сохранения высокой продукции ПНЖК в водных экосистемах. Приводятся количественные данные об оптимальных порциях потребляемой рыбы и способах её кулинарной обработки.
Введение
Почти два века назад, когда в рамках физиологии и биохимии зарождалась современная наука о питании, появился афоризм: «You are what you eat» (ты - это то, что ты ешь). Как известно, съедаемые нами органические вещества подразделяются на белки, жиры и углеводы. А ещё, как было установлено сравнительно недавно - чуть больше ста лет тому назад, в пище должны содержаться витамины. Все знают два основных свойства витаминов: 1) их требуется очень мало, если сравнивать, например, с белками; 2) большинство витаминов, как правило, не вырабатываются в организме человека и могут поступать только с особой пищей. Человеческий организм способен к биохимическому превращению поглощенной пищи и синтезу из неё многих необходимых ему для жизнедеятельности веществ. Например, мы превращаем все пищевые белки в аминокислоты, а затем из этих аминокислот строим нужные нам вещества. Кроме белков, мы можем синтезировать и жирные кислоты, но отнюдь не все. Именно поэтому в начале XX века некоторые жиры даже получили название «витамин F» (от английского Fat - жир). Но прежде чем перейти к роли «витамина F» в питании человека, кратко охарактеризуем строение и свойства жирных кислот (ЖК).
Состав и структура жирных кислот
Жиры, или липиды, - это органические вещества, практически нерастворимые в воде, но хорошо растворимые в так называемых неполярных растворителях: ацетоне, спирте, хлороформе. Основную часть липидов составляют жирные кислоты (Lehninger et al., 1993).
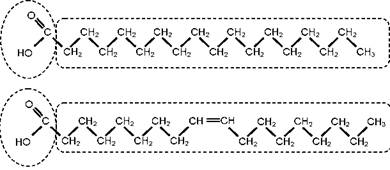
Рис . 1. Структурная формула насыщенной стеариновой кислоты 18:0 (вверху) и мононенасыщенной олеиновой кислоты 18:1n-9 (внизу)
Молекулы жирных кислот состоят из углеродной цепи, на одном конце которой находится карбоксильная (кислотная) группа (COOH), а на другом - метильная группа атомов (CH3). Разные ЖК отличаются друг от друга количеством атомов углерода, а также количеством двойных связей между атомами углерода. На рис. 1 представлено схематическое изображение двух ЖК, а на рис. 2 приведены пространственные молекулярные модели четырёх других ЖК. Следует отметить, что чем больше двойных связей в молекуле ЖК, тем сильнее закручивается углеродная цепь, приближаясь по форме к спирали (рис. 2). Пространственная структура молекул ЖК определяет их биохимические свойства, которые будут рассмотрены ниже.
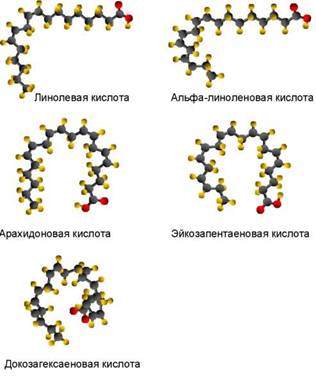
Рис. 2. Пространственные модели полиненасыщенных жирных кислот: атомы водорода, углерода и кислорода изображены жёлтыми, серыми и красными шариками соответственно
ЖК имеют номенклатурные биохимические названия, но для краткости им присвоены простые и понятные обозначения, основанные на количестве атомов углерода в цепи, а также на количестве и положении двойных связей. Например, на рис. 1 сверху изображена стеариновая (октадекановая) кислота, состоящая из 18 углеродных атомов и не имеющая двойных связей, а снизу - олеиновая (цис-9- октадеценовая), также состоящая из 18 атомов углерода, но имеющая одну двойную связь на девятом атоме углерода, если считать от ме- тильного конца молекулы. Кратко эти ЖК обозначаются как 18:0 и 18:1n-9, т. е. в начале обозначения указывается число атомов углерода (18), затем через двоеточие приводится число двойных связей (0 и 1 соответственно), а потом дан номер углеродного атома, от которого начинается двойная связь (n-9). Если двойных связей в молекуле несколько, то указывается положение первой из них. Ранее номер атома обозначался греческой буквой ю (омега), сейчас чаще используют обозначение латинской буквой n, но омега по традиции также применяется в наименовании ряда кислот, о которых речь пойдёт ниже.
ЖК, не имеющие двойных связей, называются насыщенными (стеариновая кислота 18:0 на рис. 1). ЖК с двойными связями именуются ненасыщенными (олеиновая кислота 18:1n-9 на рис. 1). Ненасыщенные кислоты, содержащие две и более двойных связи, получили специальное наименование - полиненасы- щенные жирные кислоты (ПНЖК). Именно о свойствах и физиолого-биохимической роли некоторых ПНЖК и пойдёт речь в нашей статье.
Незаменимые ПНЖК
Двойные связи в молекулу ЖК вставляют специальные ферменты - десатуразы (от англ. desaturation - уменьшение насыщенности). Каждая десатураза, представляющая собой сложную белковую молекулу, вставляет двойную связь лишь в один строго определённый участок углеродной цепи ЖК. Например, десатураза Д9 (обозначаемая прописной греческой буквой «дельта»), присоединяет двойную связь к девятому атому углерода, считаемому от карбонильного (COOH), а не от метильного конца молекулы (рис. 1). Наличие или отсутствие у разных видов организмов тех или иных десатураз определяется генотипом. Например, высшие растения и водоросли имеют гены, кодирующие десатуразы Д15 и Д12, т. е. они способны синтезировать ЖК с двойными связями в положении n-6 и n-3 (Heinz, 1993; Cohen et al., 1995; Harwood, 1996; Tocher et al., 1998). Напротив, подавляющее число видов беспозвоночных животных и все позвоночные, включая человека, этих генов не имеют и при синтезе ЖК не могут присоединять двойную связь к третьему и шестому атомам от метильного конца молекулы (Bell, Tocher, 2009; Lands, 2009).
ПНЖК, необходимые животным (и человеку), но не синтезируемые в их организмах, называют незаменимыми. К незаменимым ПНЖК относятся 18-атомные кислоты семейств n-6 и n-3 (по старому, омега-6 и омега-3): линолевая кислота с двумя двойными связями (18:2n-6) и альфа-линоленовая кислота с тремя двойными связями (18:3n-3). Ли- нолевая и альфа-линоленовая кислоты часто обозначаются аббревиатурами ЛК и АЛК соответственно. Пространственные модели ЛК и АЛК приведены на рис. 2. Животные и человек могут получать эти незаменимые ПНЖК только с пищей.
Согласно современным данным ЛК и АЛК сами по себе не играют особой роли в организме человека. 50-70 % ЛК и АЛК, поступивших с пищей, «сжигаются» для обеспечения энергетических потребностей организма в первые сутки после потребления (Broadhurst et al., 2002). Некоторые исследователи полагают, что ЛК и АЛК накапливаются в коже и содействуют её нормальному функционированию, в первую очередь предотвращают излишнюю потерю воды, а также усиливают шелушение для снижения избыточной пигментации под действием ультрафиолетового излучения (Sinclair et al., 2002).
Основная роль ЛК и АЛК в организме животных и человека состоит в том, что они могут являться биохимическими предшественниками физиологически значимых длинноцепочечных ПНЖК с 20-22 атомами углерода. Длинноцепочечные ПНЖК, называемые частично незаменимыми, - это арахи- доновая (эйкозатетраеновая) кислота (20:4n-6, АРК), эйкозапентаеновая кислота (20:5n-3, ЭПК) и докозагексаеновая кислота (22:6n-3, ДГК). Как это видно из условных обозначений, АРК относится к семейству омега-6, а ЭПК и ДГК - к семейству омега-3. Пространственные модели этих кислот приведены на рис. 2.
Как уже отмечалось, только растения имеют десатуразы Д15 и Д12 и могут синтезировать исходные ПНЖК семейства омега-6 и омега 3, т. е. линолевую и альфа-линоленовую кислоты (рис. 3). Животные, получив ЛК и АЛК с пищей, способны синтезировать из них длинноцепочечные ПНЖК омега-6 (АРК) и омега-3 (ЭПК, ДГК) (Stark et al., 2008). В синтезе участвуют ферменты, удлиняющие углеродную цепь (элонгазы), а также десату- разы Д5 и Д6 (рис. 3). Для синтеза ДГК нужен ряд дополнительных ферментов, но для простоты они не показаны на рис. 3. Однако эффективность синтеза длинноцепочечных ПНЖК у животных и человека невелика, хотя именно эти кислоты играют важнейшую роль в функционировании организма.
Роль длинноцепочечных ПНЖК в организме человека
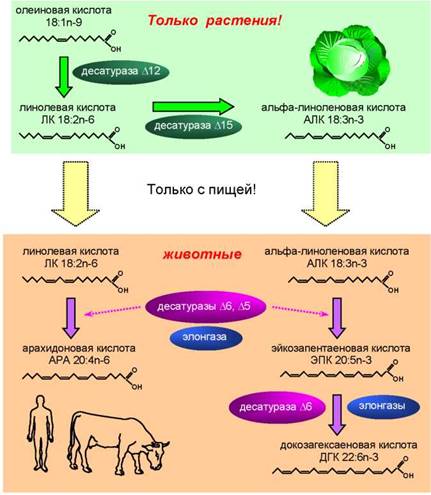
Рис. 3. Схема синтеза полиненасыщенных жирных кислот у растений и животных
Наряду с другими жирными кислотами АРК, ЭПК и ДГК входят в состав фосфолипидов клеточных мембран (Lehninger et al., 1993). Фосфолипиды обычно состоят из гидрофильной (водорастворимой) «головки» - фосфатидной кислоты и двух гидрофобных (нерастворимых в воде) «хвостов» - жирных кислот (рис. 4). Первый «хвост» присоединяется к молекуле фосфатидной кислоты в положении, обозначаемом как sn-1, и чаще всего представлен насыщенной ЖК, например стеариновой (18:0). Второй «хвост», занимающий положение sn-2, - это ненасыщенная ЖК (рис. 4). Клеточная мембрана представляет собой двойной слой (бислой) фосфолипидов со встроенными в него различными белками (рис. 5). Клеточная мембрана является основным структурно-функциональным компонентом живой клетки, и большинство процессов превращения вещества и энергии происходят именно на клеточных мембранах.
Жирнокислотный состав фосфолипидов клеток разных органов и тканей существенно различается (рис. 6). Как правило, чем сложнее функция органа, тем больше длинноцепочечных ПНЖК содержится в клетках тканей, составляющих данный орган. Например, в клетках серого вещества коры головного мозга здорового человека содержится 13 % ДГК и 9 % АРК, а содержание ДГК в сетчатке достигает 20 %, это наивысшее значение для человеческого тела (рис. 6). В то же время в ади- позной (жировой) ткани, которая состоит не из фосфолипидов, а из запасных жиров - тригли- церинов, содержится менее 1 % ДГК (рис. 6).
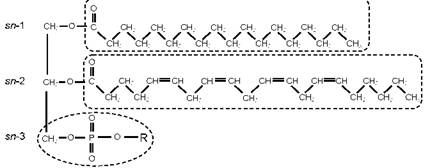
Рис. 4. Структурная формула молекулы фосфолипида: в положении sn-1 изображена стеариновая кислота (как на рис. 1), а в положении sn-2 - длинноцепочечная полиненасыщенная арахидоновая кислота (АРК, 20:4n-6). Головная группа R в положении sn-3 может быть представлена атомом водорода, спиртами и некоторыми другими веществами
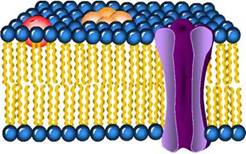
Рис. 5. Пространственная модель клеточной мембраны. Фосфолипиды имеют синюю «головку» и два жирнокислотных «хвоста». Также в мембрану включены белковые глобулы
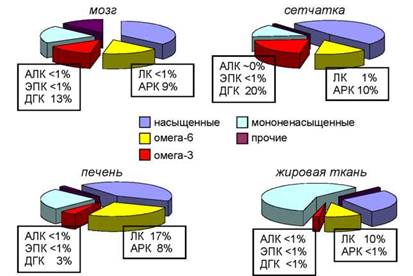
Рис. 6. Содержание ( % от суммы ЖК) различных групп жирных кислот в клетках тканей человека (Lauritzen et al., 2001; McNamara et al., 2006)
Таким образом, ДГК является основной полиненасыщенной жирной кислотой в клеточных мембранах сетчатки глаза (в фоторецепторах), а также в нервных клетках. Считается, что благодаря своей длинной цепи (22 атома) и шести двойным связям ДГК имеет уникальную стереохимическую пространственную структуру: она почти закручена в спираль (рис. 2), и именно эта молекула в составе специализированных клеточных мембран обеспечивает наиболее эффективное восприятие светового сигнала и проведение нервного импульса (SanGiovanni, Chew, 2005).
Важнейшая физиолого-биохимическая роль двух других длинноцепочечных ПНЖК, АРК и ЭПК, состоит в том, что они биохимические предшественники синтеза эндогормонов - эйкозаноидов (SanGiovanni, Chew, 2005). Синтез эндогормонов (рис. 7) начинается с высвобождения ПНЖК из фосфолипидов клеточных мембран под действием особого фермента - фосфолипазы А2 (обозначаемой латинскими буквами PLA2). Эта фосфоли- паза A2 способна отщеплять именно ПНЖК, находящуюся в молекуле фосфолипида в положении sn-2 (рис. 4). Затем другие ферменты, циклооксигеназы (COX), синтезируют из свободных ПНЖК простагландины (PG) и тромбоксаны (TX), а липоксигеназы (LOX) синтезируют лейкотриены (LT) (рис. 7). Важно отметить, что из арахидоновой кислоты синтезируются в основном простагландины и тромбоксаны так называемой второй серии, т.е. имеющие две двойные связи и обозначаемые PG-2 и TX-2 соответственно, а также лейкотриены четвёртой серии LT-4. TX-2 вызывают сужение кровеносных сосудов, усиливают агрегацию (слипание) тромбоцитов. Чрезмерная агрегация тромбоцитов приводит к повышению артериального давления, образованию тромбов и закупорке сосудов (рис. 7). PG-2 запускают воспалительный процесс и индуцируют боль. LT-4 вызывают спазмы бронхов и секрецию слизи (рис. 7). Из эйкозапентаеновой кислоты получаются эндогормоны, обладающие противоположными свойствами, чем производные АРК (Simopoulos, 2000). Из ЭПК синтезируются простагландины и тромбоксаны третьей серии (с тремя двойными связями), PG-3 и TX-3, и лейкотриены пятой серии LT-5 (рис. 7). TX-3 вызывают расширение кровеносных сосудов, препятствуют слипанию тромбоцитов и тем самым снижают артериальное давление. PG-3 обладают противовоспалительным эффектом (Wall et al., 2010), а LT-5 являются антиаллергенами и расширяют бронхи (рис. 7).
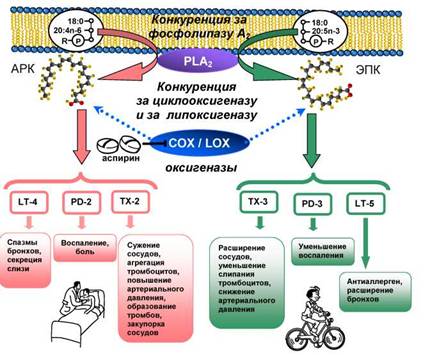
Рис. 7. Схема синтеза эндогормонов — эйкозаноидов в организме человека (пояснения в тексте)
Следует подчеркнуть, что синтез из АРК и ЭПК эндогормонов, обладающих противоположным действием на организм, обеспечивается одними и теми же ферментами: фос- фолипазой A2 и циклооксигеназами (рис. 7). Таким образом, если в фосфолипидах клеток животных и человека имеется избыток АРК, то ферменты быстро превращают их в проста- гландины, тромбоксаны и лейкотриены PG- 2, TX-2 и LT-4, избыточный синтез которых приводит к опасным заболеваниям, прежде всего сердечно-сосудистым, к воспалению, отекам, аллергии и боли. Конечно, против этих болезней и симптомов имеется целый ряд лекарств. Например, всем известный аспирин блокирует циклооксигеназу (рис. 7). Но всех перечисленных выше неприятностей можно избежать, если в фосфолипидах содержится достаточное количество ЭПК, которая конкурирует с АРК за ферменты PLA2 и COX/ LOX (рис. 7). Фосфолипаза и циклооксиге- назы, «отвоёванные» у АРК, производят из ЭПК благоприятные для здорового организма эндогормоны PG-3, TX-3 и LT-5 (рис. 7). Следовательно, чтобы боль и воспаление не умерщвляли, а излечивали, необходим определённый баланс эндогормонов - производных АРК и ЭПК в организме.
Необходимо добавить, что ДГК под действием фосфолипазы и циклооксигеназы может также превращаться в эндогормон - до- козаноид, называемый нейропротектином D (Bazan, 2009). Уже из названия этого эндогормона ясна его функция: защита нервных клеток от повреждения, например от окислительного стресса.
Перечисленные выше биохимические механизмы действия каждой отдельной ПНЖК в организме человека были открыты сравнительно недавно: менее 40 лет назад (Plourde, Cunnane, 2007). А 70 лет назад, когда было эмпирически установлено, что нормальный рост и развитие животных невозможны без жирных кислот омега-6 и омега-3, все эти кислоты обозначали как «витамин F», поскольку особая роль каждой из них не была известна. Да и надёжные методы, позволяющие идентифицировать ПНЖК в биологических объектах, т.е. отличать одну кислоту от другой в их смеси, получили широкое распространение ещё позднее - в середине 90-х годов прошлого века, т.е. менее 20 лет назад. Таким методом является современная хроматомасс- спектрометрия с использованием капиллярных колонок (рис. 8).
Медицинские исследования ПНЖК

Рис. 8. Современный хроматомасс-спектрометр позволяет специалистам точно определять содержание отдельных ПНЖК в любых биологических объектах
После расшифровки механизмов действия ПНЖК в организме начался период их массовых клинических и эпидемиологических исследований. В первую очередь изучали связь между содержанием ПНЖК в плазме крови и наличием сердечно-сосудистых заболеваний. Во второй половине XX века смертность от сердечно-сосудистых заболеваний в индустриально развитых западных странах начала угрожающе расти и вышла на первое место среди смертности от всех остальных заболеваний. Например, в России в 1995-2009 годах ежегодно от болезней системы кровообращения умирали около 1 млн 200 тыс. чел. тогда как от внешних причин (убийства, самоубийства, отравление алкоголем, транспортные происшествия и т.д.) - около 300 тыс. чел., и от раковых заболеваний - также около 300 тыс. чел. (Попов, 2012). Таким образом, смертность от сердечно-сосудистых заболеваний в России составляла в последние два десятилетия более 55 % от всей смертности (Попов, 2012). К сожалению, по этому печальному показателю наша страна занимает первое место в мире. Сердечно-сосудистые заболевания уже давно связывают с содержанием липидов в крови. Если ранее медики обращали внимание на содержание «витамина F» - общей суммы ПНЖК (ЛК, АЛК, АРК и др.), то в последние десятилетия, в связи с обнаружением различий в физиологобиохимических функциях омега-6 и омега-3 кислот, исследовалась специфическая роль каждой из этих групп.
Ещё в середине 1970-х годов было обнаружено, что в плазме крови у гренландских эскимосов, среди которых сердечно-сосудистые заболевания почти отсутствуют, содержится значительно меньше омега-6 кислот (ЛК, АРК) и значительно больше омега-3 ПНЖК (ЭПК, ДГК), чем у населения стран Западной Европы (Wall et al., 2010). А вот содержание холестерина, ранее считавшегося главным фактором риска, в крови у эскимосов и европейцев было практически одинаковым. Дальнейшие клинические и эпидемиологические (популяционные) исследования в основном проводили в североамериканских и западноевропейских странах на очень больших группах пациентов, некоторые включали более десяти тысяч человек. Эти исследования доказали, что повышенное потребление омега-3 ПНЖК достоверно (почти в 10 раз!) снижает риск сердечно-сосудистых заболеваний у здоровых людей, способствует выздоровлению и на 35 % снижает смертность среди людей, перенесших эти заболевания (Harris et al., 2009). Очевидно, механизм благоприятного влияния ЭПК на функционирование кровеносной системы заключается в повышении синтеза эйкозаноидов, расширяющих сосуды, снижающих тромбообразование, артериальное давление и воспаление (Plourde, Cunnane, 2007; Phang et al., 2011). Полезное воздействие ДГК, вероятно, состоит в обеспечении эффективного проведения сигналов в нервных клетках, препятствующих аритмии и спазмам сердца и сосудов (Plourde, Cunnane, 2007; Phang et al., 2011). Высокий уровень ДГК в мембранах митохондрий (клеточных «генераторов энергии») сердечной мышцы повышает эффективность производства и использования энергии сердцем (SanGiovanni, Chew, 2005). Хотя не ясно, какой из этих механизмов является ведущим, необходимость длинноцепочечных омега-3 ПНЖК, ЭПК и ДГК для поддержания здоровья сердечно-сосудистой системы - доказанный медицинский факт (Plourde, Cunnane, 2007). В настоящее время для определения риска сердечно-сосудистых заболеваний предложен омега-3 индекс, представляющий собой процент ЭПК+ДГК от суммы ЖК в клетках красной крови (эритроцитах). У пациентов с омега-3 индексом <4 % риск этих заболеваний в 10 раз выше, чем у пациентов с индексом >8 % (Saldanha et al., 2009).
Сбалансированная диета как способ профилактики сердечно-сосудистых заболеваний
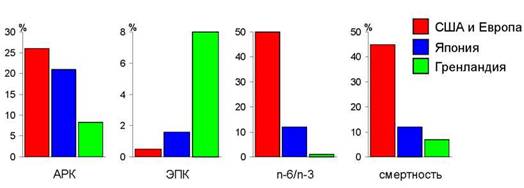
Рис . 9. Содержание ( % от суммы ЖК) арахидоновой и эйкозапентаеновой кислот в фосфолипидах тромбоцитов (клеток крови) населения разных стран, соотношение у них n-6 и n-3 ПНЖК, и доля ( %) смертности от сердечно-сосудистых заболеваний в общей смертности в этих странах (по Simopoulos, 2000)
Итак, из современных биохимических данных следует, что в организме человека, во-первых, должно содержаться достаточное количество омега-3 ПНЖК. Во-вторых, соотношение омега-6 и омега-3 кислот тоже имеет важнейшее значение для кровеносной системы. Эти показатели тесно связаны со смертностью от сердечно-сосудистых заболеваний. Например, у населения США и Европы в клетках крови (тромбоцитах) содержание арахидоновой n-6 кислоты почти в три раза выше, а содержание n-3 эйкозапентаено- вой кислоты в 16 раз ниже, чем у эскимосов Гренландии (рис. 9). Соотношение n-6:n-3 у населения этих стран различается в 50 раз, и смертность от сердечно-сосудистых заболеваний в США и Европе почти в 7 раз выше, чем в Гренландии (рис. 9). Население Японии, занимающее по показателям количества и соотношения n-6 и n-3 ПНЖК в крови промежуточное положение, имеет и промежуточное значение смертности от сердечно-сосудистых заболеваний: около 12 % от общей смертности по сравнению с 45 % в США и 7 % в Гренландии (рис. 9).
Уровень различных ПНЖК в крови и других тканях и органах человека напрямую зависит от его пищи. На основании многолетних клинических исследований и эпидемиологических наблюдений, охвативших несколько сотен тысяч человек, Всемирная организация здравоохранения и ряд национальных медицинских организаций рекомендовали для предотвращения сердечно-сосудистых заболеваний ежедневное потребление 5001000 мг ЭПК+ДГК на человека (Kris-Etherton et al., 2002, 2009; Reis, Hibbeln, 2006; Harris et al., 2009). При этом соотношение потребляемых n-6 и n-3 ПНЖК, согласно рекомендациям Национального института здоровья США и японских национальных фондов, должно быть не ыше 2:1-3:1 (Davis, Kris-Etherton, 2003).
Однако проблема состоит в том, что в современных обществах так называемого западного типа, т.е. в большинстве индустриально развитых стран, соотношение n-6:n-3 в продуктах питания составляет в настоящее время 15:1 - 25:1 (Simopoulos, 2000; Wall et al., 2010). Этот показатель начал существенно увеличиваться со второй половины XX века в связи с модернизацией сельского хозяйства и преобладанием мясной продукции, выращиваемой на кормах, богатых зерном с высоким содержанием омега-6 ПНЖК (Simopoulos, 2000). Тенденция увеличения n-6:n-3 в продуктах питания продолжается до сих пор. Например, в Европе потребление n-6 линолевой кислоты за последние двадцать лет возросло на 50 % (Wall et al., 2010). Одновременно с ростом соотношения n-6:n-3 в пище наблюдается рост сердечнососудистых заболеваний. Хотя понятно, что увеличение соотношения n-6:n-3 в пище не единственный фактор, вызывающий болезни органов кровообращения, в настоящее время имеются все основания полагать, что его роль достаточно велика.
Таблица 1. Содержание ( % от суммы жирных кислот) линолевой (ЛК, 18:2n-6), альфа-линоленовой (АЛК, 18:3n-3), арахидоновой (АРК, 20:4n-6), эйкозапентаеновой (ЭПК, 20:5n-3) и докозагексаеновой (ДГК, 22:6n-3) кислот в различных продуктах питания и соотношение в них сумм n-6 и n-3 ПНЖК
| Продукт |
ЛК |
АЛК |
АРК |
ЭПК |
ДГК |
n-6/n-3 |
Источник |
| Льняное масло |
16.0 |
57.0 |
0.0 |
0.0 |
0.0 |
0.3 |
Davis, Kris-Etherton, 2003 |
| Рапсовое масло |
21.0 |
11.0 |
0.0 |
0.0 |
0.0 |
2 |
Davis, Kris-Etherton, 2003 |
| Соевое масло |
51.0 |
7.0 |
0.0 |
0.0 |
0.0 |
7 |
Davis, Kris-Etherton, 2003 |
| Оливковое масло |
5.4 |
0.6 |
0.0 |
0.0 |
0.0 |
9 |
Rodriguez-Carpena et al., 2012 |
| Подсолнечное масло |
62.0 |
0.3 |
0.0 |
0.0 |
0.0 |
222 |
Rodriguez-Carpena et al., 2012 |
| Грецкие орехи |
58.0 |
14.0 |
0.0 |
0.0 |
0.0 |
4 |
Davis, Kris-Etherton, 2003 |
| Пшеница |
51.9 |
4.0 |
0.0 |
0.0 |
0.0 |
13 |
Suchowilska et al., 2009 |
| Капуста |
19.6 |
60.6 |
0.3 |
0.0 |
0.0 |
0.3 |
Batista et al., 2011 |
| Свинина |
14.2 |
0.9 |
2.2 |
0.3 |
0.0 |
7.2 |
Wood et al., 2008 |
| Баранина |
2.7 |
1.4 |
0.6 |
0.4 |
0.0 |
1.3 |
Wood et al., 2008 |
| Говядина |
2.4 |
0.7 |
0.6 |
0.3 |
0.0 |
2.1 |
Wood et al., 2008 |
| Жареный гамбургер (на подсолнечном масле) |
34.8 |
0.6 |
0.7 |
0.7 |
0.01 |
20 |
Rodriguez-Carpena et al., 2012 |
| Курятина |
17.0 |
0.4 |
4.7 |
0.2 |
0.3 |
18.5 |
Rule et al., 2002 |
| Куриные яйца |
26.1 |
0.5 |
5.0 |
0.0 |
1.1 |
19.9 |
Simopoulos, 2000 |
| Треска |
1.4 |
2.0 |
2.8 |
7.6 |
50.8 |
0.1 |
Usydus et al., 2011 |
| Сельдь |
4.4 |
5.3 |
0.7 |
6.2 |
20.4 |
0.2 |
Usydus et al., 2011 |
| Форель |
6.5 |
7.2 |
0.5 |
8.0 |
17.5 |
0.2 |
Usydus et al., 2011 |
| Карп |
7.3 |
5.9 |
1.4 |
2.3 |
2.1 |
0.9 |
Usydus et al., 2011 |
Виды продуктов с высоким уровнем тех или иных ПНЖК перечислены в табл. 1. Как следует из приведённых данных, в подсолнечном масле почти нет альфа-линоленовой кислоты и для него характерно очень высокое соотношение n-6:n-3 кислот. В оливковом масле данное соотношение диетически более благо - приятно, но это масло вытесняется с рынка более дешёвым подсолнечным, и в последнее десятилетие даже в таких средиземноморских странах, как Испания и Португалия, подсолнечного масла производится больше, чем оливкового (Sanders, 2000). Неблагоприятно высокие соотношения n-6:n-3 имеются в пшенице, а также в курином мясе и куриных яйцах, если этих птиц кормят зерном (табл. 1). В мясе, особенно в баранине и говядине, соотношение омега-6 и омега-3 кислот фактически идеально, и даже в свинине оно относительно невелико. Однако если мясо пожарить на подсолнечном масле, то n-6:n-3 резко увеличится до 20, как, например, в популярном в западных странах гамбургере (табл. 1). Поскольку АЛК - основная жирная кислота фотосинтезирующих мембран хлоропластов, её особенно много в зелёных листьях и других органах растений: в капусте, в салате и др. Различные рыбы также богаты омега-3 кислотами, особенно длинноцепочечными, ЭПК и ДГК (табл. 1). Таким образом, для достижения благоприятного соотношения n-6:n-3 в пище, снижающего риск сердечно-сосудистых заболеваний, необходимо употреблять больше зелёных растений и рыбы. Мясо животных само по себе не является «опасным» продуктом, но на соотношение в нём омега-6 и омега-3 ПНЖК влияет способ кулинарной обработки (табл. 1).
Рыба - основной источник длинноцепочечных ПНЖК для человека
Как отмечалось выше, 18-атомная АЛК, чрезвычайно важная для растений, в организме животных не играет самостоятельной роли, но является предшественником для синтеза физиологически значимых длинноцепочечных кислот, ЭПК и ДГК (рис. 3). Если у травоядных животных потребности в ЭПК и ДГК, вероятно, могут практически полно - стью обеспечиваться их синтезом из альфа- линоленовой кислоты поедаемых зелёных растений, то для большинства всеядных и хищников, включая человека, по современным данным, необходимо прямое потребление длинноцепочечных ПНЖК. У большинства людей со среднестатистическим генотипом способность к синтезу ЭПК и ДГК из АЛК весьма невелика и не обеспечивает физиологических потребностей организма. Более 60 % АЛК, поступившей с пищей, в первые же 8 часов «сжигаются» в митохондриях в процессе бета-окисления, т.е. тратятся на производство энергии (Plourde, Cunnane, 2007). Для сравнения: менее 5 % потреблённой ДГК идёт на бета-окисление, тогда как оставшаяся основная часть встраивается в клеточные мембраны. Согласно современным данным в среднем лишь около 10 % пищевой АЛК может быть конвертировано в ЭПК и лишь около 5 % - в ДГК (Davis, Kris-Etherton, 2003; Wall et al., 2010). Понятно, что увеличение АЛК в пище не всегда сопровождается увеличением её производных — ЭПК и ДГК в крови. Вероятно, именно поэтому потребление с пищей повышенных количеств АЛК не всегда даёт отчётливый клинический эффект. А вот прямое потребление этих физиологически важных длинноцепочечных ПНЖК вызывает пропорциональное увеличение их концентрации в тканях организма человека (Hibbeln et al., 2006). Как уже говорилось, достоверно доказано, что употребление оптимальной дозы — около 1 г ЭПК+ДГК в сутки — способствует существенному улучшению работы органов кровообращения и нервной системы и в значительной степени помогает предотвратить соответствующие заболевания.
Таким образом, большинству людей для борьбы с истинной чумой XXI века — сердечно-сосудистыми заболеваниями — следует употреблять рекомендованные современной наукой дозы ЭПК+ДГК. Конечно, есть относительно небольшие группы людей, например вегетарианцы, которые могут обходиться без пищевых длинноцепочечных ПНЖК. Уровень ЭПК и ДГК в крови вегетарианцев на 20-30 % ниже, чем у «всеядных» людей, однако у них нет явно выраженных клинических симптомов недостатка ПНЖК (Davis, Kris-Etherton, 2003). Причины подобных особенностей организма до концы не ясны, однако в целом понятно, что в тканях таких людей ЭПК и ДГК должны более эффективно синтезироваться из растительной АЛК и экономнее расходоваться (Plourde, Cunnane, 2007). Ничего удивительного в этом нет, поскольку отличия в работе ферментных систем — в данном случае Д5 и Д6 десатураз — у людей с разным генотипом хорошо известны. К тому же вегетарианцы употребляют с растительной пищей сравнительно большое количество АЛК — исходной кислоты для последующего синтеза длинноцепочечных ПНЖК омега-3 — и не получают с мясом готовой омега-6 АРК, конкурирующей с ЭПК за фосфолипазу А2 при синтезе эндогормонов (рис. 7). Вероятно, этот генетически обусловленный способ регуляции и обеспечивает нормальное функционирование организма.
Итак, для основной массы людей со среднестатистическим генотипом необходимо регулярное употребление значительных количеств ЭПК и ДГК. Из табл. 1 видно, что основным продуктом, содержащим высокие концентрации этих длинноцепочечных омега-3 ПНЖК, является рыба. Почему именно рыба и другие морепродукты — крабы, моллюски, креветки — так богаты ЭПК и ДГК? Наземные высшие (цветковые) растения останавливают свой синтез на 18-атомной альфа-линоленовой кислоте (рис. 3) и не синтезируют длинноцепочечные омега-3 ПНЖК (Heinz, 1993; Tocher et al., 1998). Как отмечалось выше, большинство животных обладают слабой способностью конвертировать АЛК в ЭПК и ДГК. Из всех известных организмов лишь некоторые микроводоросли (диатомеи, перидинеи, криптофиты) способны эффективно синтезировать и накапливать в своей биомассе большие количества ЭПК и ДГК. То есть водные экосистемы — озёра, реки и моря — являются основными месторождениями длинноцепочечных омега-3 ПНЖК (Gladyshev et al., 2009a). ЭПК и ДГК, синтезированные микроводорослями, по трофической (пищевой) цепи передаются к водным беспозвоночным, от них — к рыбам и затем — к человеку и другим наземным животным (рис. 10).
Вероятная роль потребления рыбы в эволюции человека
Не исключено, что поток ПНЖК из водных экосистем в наземные стал важнейшим фактором в эволюции человека. Как отмечалось выше, ДГК является основной жирной кислотой в мембранах клеток серого вещества коры головного мозга человека. Человек отличается от всех остальных животных именно размером и массой головного мозга. Связь размеров мозга в сравнении с размерами тела описывается так называемым коэффициентом энцефализации (от греч. en cephalos - головной мозг, «находящийся внутри головы») - это отклонение истинных размеров мозга от соотношения, рассчитанного по «стандартному» виду млекопитающих (Roth, Dicke, 2005). Как видно из рис. 11, коэффициент энцефализации у современного Homo sapiens намного выше, чем у австралопитека и человекообразных обезьян, не говоря уж о других млекопитающих. Сухое вещество мозга на 60 % состоит из липидов (Broadhurst et al., 2002), из этих липидов 35 % составляют жирные кислоты (Lauritzen et al., 2001), среди которых наибольшая доля (до 20 %) принадлежит ДГК (McNamara, Carlson, 2006). При этом важно отметить, что содержание ДГК в мозге всех млекопитающих является почти одинаковым
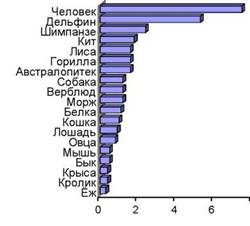
Коэффициент энцефализации
Рис. 11. Величина коэффициента энцефализации у человека и других млекопитающих (Roth, Dicke, 2005)
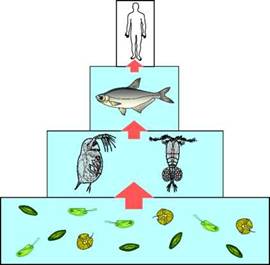
Рис. 10. Пищевая цепь, по которой. человеку передаются длинноцепочечные омега-3 ПНЖК, синтезированные диатомовыми, криптофитовыми и перидиниевыми микроводорослями
(Broadhurst et al., 2002). Поскольку в самом мозге ДГК почти не синтезируется, высокий коэффициент энцефализации означает, что организм человека должен снабжать свой мозг ДГК гораздо интенсивнее, чем организм всех остальных видов животных.
В период интенсивного формирования мозга во время внутриутробного развития человеческий плод получает ДГК из организма матери. При этом плацента избирательно поглощает материнскую ДГК и передаёт эту ПНЖК плоду. Например, скорость переноса ДГК через плаценту в три раза выше, чем АРК (Lauritzen et al., 2001). В связи с интенсивным и избирательным переносом через плаценту содержание ДГК в плазме крови матери снижается в два раза (Broadhurst et al., 2002). Схожее явление обнаружено нами у рыб: во время вынашивания икры, имеющей чрезвычайно высокий уровень накопления ДГК, содержание данной кислоты в мышцах рыбы уменьшалось почти в два раза (Sushchik et al., 2007). В период грудного вскармливания ребёнка запасы ДГК в организме матери также продолжают истощаться, поскольку эта ПНЖК поступает в грудное молоко (Lauritzen et al., 2001). ДГК из крови избирательно поглощается именно клетками мозга, а также нервной системы и органов зрения (Bazan, . Клетки этих органов способны чрезвычайно долгое время удерживать «захваченную» ДГК, обеспечивая её постоянную концентрацию. Например, чтобы добиться снижения содержания ДГК в мозге и сетчатке у грызунов, необходимо держать на диете без ДГК два их поколения (Bazan, 2009). Считается, что столь же эффективная консервация ДГК характерна и для мозга человека (Lauritzen et al., 2001). Однако, по некоторым оценкам, в мозге человека в результате метаболизма ежесуточно расходуется 2-8 % ДГК, и эти потери должны восполняться организмом (McNamara, Carlson, 2006).
Недостаток ДГК в диете матери и ребёнка приводит к снижению способностей к обучению, зрительной активности, психомоторных функций детей (McNamara, Carlson, 2006; Reis, Hibbeln, 2006). Дефицит ДГК у взрослых вызывает повышенный риск депрессии, шизофрении, агрессии, слабоумия и прочих нервных расстройств, включая болезнь Альцгеймера (Davis, Kris-Etherton, 2003; Hibbeln et al., 2006; Robert, 2006; Plourde, Cunnane, 2007; Saldanha et al., 2009). Для профилактики нервных расстройств и психических заболеваний Американская психиатрическая ассоциация рекомендует ежедневное потребление не менее 1 г омега-3 ПНЖК (Reis, Hibbeln, 2006). Следует отметить, что, хотя объём данных клинических и эпидемиологических исследований о пользе ЭПК+ДГК для предотвращения и лечения нервных и психических заболеваний постоянно растёт, рекомендуемые дозы потребления ПНЖК остаются такими же, как и рекомендуемые для профилактики сердечно-сосудистых заболеваний (Harris et al., 2009). Необходимо также подчеркнуть, что в настоящее время нет средств для лечения одного из самых опасных и распространённых нервных заболеваний - болезни Альцгеймера (Harris et al., 2009; Wall et al., , однако появляются обнадёживающие данные, свидетельствующие о возможности снизить риск этого заболевания, употребляя ДГК (Wall et al., 2010).
Как уже говорилось, ДГК в организме большинства людей лишь в очень небольших количествах синтезируется из АЛК, полученной из растительной пищи, поэтому очень важно употреблять в пищу продукты, богатые ДГК, а именно рыбу (Philibert et al., 2006; Plourde, Cunnane, 2007). В последние два десятилетия группа учёных развивает теорию об уникальной роли рыбной пищи в эволюции человека (Crawford et al., 1999; Broadhurst et al., 2002; Muskiet et al., 2004). Данная теория базируется на двух фактах, упомянутых выше. Во-первых, для развития уникально большого (относительно размеров тела) мозга человеку требуется сравнительно большое количество ДГК. Во-вторых, рыба и другие водные организмы являются единственной пищей, содержащей значительные количества ДГК. Первобытный человек, как известно, селился на берегах рек, озёр и морей. Наверняка тому благоприятствовало множество причин. Доступ к пище, исключительно богатой ДГК, необходимой для эволюции мозга, действительно мог стать одним из важных факторов, благодаря которому первобытные люди стали отличаться от приматов, обитавших в глубине лесов или саванн. Рыба, оставшаяся на берегу моря в так называемых приливных лужах, моллюски, черепахи могли стать относительно лёгкой добычей даже безоружных людей (Gibbons, 2002). Причём эту добычу могли собирать женщины и дети, т.е. особи, наиболее нуждающиеся в ДГК. Затем взрослые мужчины, накопившие изрядный запас ДГК в нервной и зрительной ткани, могли отправляться вглубь континента на охоту за крупными наземными животными. Интенсивное питание людей каменного века морепродуктами имеет археологические подтверждения. Например, в пещерных стоянках в устье реки Класиес на побережье Южной Африки ископаемые слои раковин моллюсков, съеденных человеком, достигают толщины 20 метров (Broadhurst et al., 2002).
Водные экосистемы - основной производитель длинноцепочечных ПНЖК
В связи с недавно определённой ролью водных экосистем как основного источника длинноцепочечных омега-3 ПНЖК в нашей биосфере возникло три вопроса.
Вопрос первый: человек и некоторые животные, например медведи, - умелые рыболовы; а каким образом получают водные ПНЖК другие наземные животные, не способные ловить рыбу?
Вопрос второй: каковы потери ПНЖК в пищевой цепи? Как известно, согласно закону экологической пирамиды биомасса организмов каждого последующего трофического уровня составляет лишь около 10 % биомассы предыдущего уровня (рис. 10). То есть между каждой парой звеньев трофической цепи сжигается 90 % передаваемого по этой цепи органического вещества. Как же ПНЖК вообще доходят до наземных хищников и человека?
Вопрос третий: хватает ли ПНЖК, синтезируемых в водных экосистемах, для обеспечения всех нуждающихся в них наземных животных и человека?
Наземные животные, не умеющие ловить рыбу, могут получать ПНЖК из водных экосистем двумя путями (Gladyshev et al., 2009a). Первый поток «организуют» околоводные птицы, питающиеся водными организмами - водорослями, мелкими беспозвоночными и той же рыбой. «Доставив» выловленные в воде ПНЖК на сушу в своей биомассе, эти птицы, их яйца или птенцы становятся добычей наземных хищников (рис. 12А). Согласно глобальным расчётам, т.е. сделанным для всей биосферы в целом, околоводные птицы в год выносят на сушу приблизительно 432 тыс. т ЭПК+ДГК (рис. 12А). Для сравнения: все медведи, охотящиеся на идущих на нерест в реки тихоокеанских лососей, в год добывают лишь 2 тыс. т ЭПК+ДГК (рис. 12В). Важно отметить, что вклад Мирового океана в общий поток ПНЖК из воды на сушу оказался в 25 раз ниже, чем вклад континентальных водоёмов - озёр и рек. Этот удивительный на первый взгляд факт объясняется очень просто: для расчёта потоков важна не площадь водоёма, а его периметр (длина береговой линии), то есть длина границы контакта вода-суша. Глобальная длина береговой линии океана составляет 594 000 км, тогда как суммарная береговая линия сотен миллионов малых и больших озёр имеет длину около 35 млн км, т.е. почти в 60 раз превышает пери- метр океана (Gladyshevetal., 2009a). Следует подчеркнуть, что все рассуждения о длине береговой линии и ведущей роли внутренних водоёмов в снабжении наземных экосистем незаменимыми ПНЖК касаются только природных сообществ. Человек, вооружённый современными техническими средствами, добывает основное количество ПНЖК из океана (речь об этом пойдёт ниже).
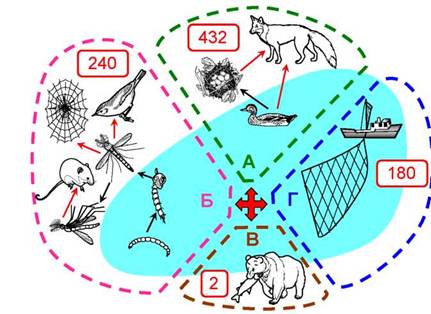
Рис. 12. Потоки ПНЖК (ЭПК+ДГК) из воды на сушу: А - за счёт водных птиц (432 тыс. т в год); Б - за счёт вылета амфибионтных насекомых (240 тыс. т в год); В - за счёт питания наземных хищников (2 тыс. т в год); Г - вылов человеком (180 тыс. т в год) (Gladyshev et al., 2009)
Другой существенный поток длинноцепочечных омега-3 ПНЖК из водных экосистем на сушу осуществляется за счёт вылета амфибионтных насекомых (рис. 12Б). Амфи- бионтными называются те летающие насекомые, личинки и куколки которых растут и развиваются в воде: стрекозы, мошки, ручейники, кровососущие комары, комары-звонцы и др. Водные личинки питаются водорослями и другими мелкими организмами, запасают ПНЖК, окукливаются, и когда из куколки вылетает взрослое насекомое (имаго), оно несёт в своём теле на сушу эти продукты синтеза микроводорослей (рис. 12Б). Глобальный вынос ЭПК+ДГК из воды на сушу амфибионт- ными насекомыми составляет приблизительно 240 тыс. т в год (Gladyshev et al., 2009a). Таким образом, не вдаваясь в детали, которые ещё долго предстоит уточнять, мы уже можем в общем виде ответить на первый вопрос.
Что же касается второго вопроса о потерях ПНЖК в пищевой цепи (трофической пирамиде, рис. 10), то ответ на него также удалось получить несколько лет назад (Gladyshev et al., 2011b). В небольшом водохранилище параллельно определялась эффективность переноса по трофической цепи общего органического углерода, ЭПК и ДГК, а также других полиненасыщенных жирных кислот с 16 атомами углерода, которые не используются животными для построения клеточных мембран. Эффективность переноса углерода оказалась близка к классической величине и составила чуть более 6 %. 16-атомные ПНЖК служили прекрасным «топливом» для клеток и «сжигались» в митохондриях почти на 95 %, т.е. они переносились между трофическими уровнями с эффективностью 5 %. А вот эффективность переноса омега-3 ПНЖК составила около 13 %, т.е. была в два раза выше, чем эффективность переноса всех суммарных органических веществ (углерода). Таким образом, было доказано, что омега-3 ПНЖК, производимые микроводорослями, не «сжигаются», а накапливаются в биомассе организмов верхних трофических уровней (Gladyshev et al., 2011b). Поэтому в съедаемой нами рыбе содержание ЭПК+ДГК существенно выше, чем в биомассе водорослей, которые изначально синтезируют эти кислоты.
Следует отметить, что высокая эффективность переноса длинноцепочечных омега-3 ПНЖК в пищевой цепи и их накопление в биомассе верхних трофических звеньев экосистемы имеют некий аналог и на организ- менном уровне. Напомним, что мозг человека и других животных избирательно поглощает ДГК из крови и эта жирная кислота накапливается и сохраняется в его клетках (Bazan, . Эффективность переноса ДГК плацентой человека к плоду существенно выше, чем эффективности переноса других жирных кислот (Lauritzen et al., 2001). ДГК накапливается и в икре рыб за счёт снижения её содержания в мышечной ткани (Sushchik et al., 2007).
Ответ на третий вопрос представляет большую сложность, поскольку в различных климатических поясах и ландшафтах функционируют различные водные и наземные экосистемы и в идеале для каждого ландшафта должно быть определено соотношение продукции ПНЖК и потребности в них. Однако приблизительные расчёты можно сделать, опираясь на средние для биосферы величины. Для таких приблизительных расчётов было использовано среднее для нашей планеты значение потока ПНЖК из континентальных водоёмов на сушу, а в качестве «подопытных» животных были взяты всеядные грызуны, биомасса которых в наземных экосистемах в десятки раз превышает биомассу крупных хищников (Gladyshev et al., 2009a). Выяснилось, что в среднем поток ЭПК+ДГК на единицу площади наземных экосистем составляет от 2,5 до 11,8 кг на 1км2 в год, а потребности всеядных грызунов оцениваются в кг на 1км2 в год. Таким образом, в среднем продукции длинноцепочечных омега-3 ПНЖК хватает, чтобы обеспечить потребности даже самых активных и многочисленных наземных животных, но всё же крайнее значение этой продукции может быть и ниже «прожиточного минимума», тем более что обычно в экосистемах потребляется не вся продукция, а лишь непосредственно доступная консументам.
Согласно расчётам (Gladyshev et al., 2009a), опирающимся на мировые данные о величинах уловов рыбы и беспозвоночных и на средние значения содержания в их биомассе длинноцепочечных ПНЖК, человек ежегодно вылавливает из водных экосистем 180 тыс. т ЭПК+ДГК (рис. 12Г). Это почти в раза меньше, чем птицы (рис. 12А), но в 90 раз больше, чем медведи (рис. 12В).
По данным ООН, каждый человек на планете потребляет в среднем 16 кг рыбы и морепродуктов в год, включая аквакультуру. Среднее содержание ЭПК+ДГК в биомассе рыб и беспозвоночных составляет 2 мг на 1 г (Gladyshev et al., 2009a). Отсюда легко подсчитать, что ежесуточное среднее потребление ЭПК+ДГК человеком составляет около 0,1 г. Напомним, что в соответствии с современными рекомендациями международных и национальных медицинских организаций для предотвращения сердечно-сосудистых заболеваний требуется ежесуточное потребление около 1 г ЭПК+ДГК. Отсюда следует неутешительный и тревожный вывод: человечество испытывает большой дефицит физиологически важных длинноцепочечных омега-3 ПНЖК, который способствует чрезвычайно высокой смертности от болезней органов кровообращения. Особенно остро данная проблема стоит в России.
Как сохранить и приумножить продукцию необходимых для человека ПНЖК в водных экосистемах
Итак, согласно медицинским предписаниям для предотвращения сердечно-сосудистых заболеваний, являющихся главной причиной смертности населения во многих странах, необходимо в несколько раз увеличить потребление ПНЖК. Однако, по мнению экспертов (Pauly et al., 2002), вылов рыбы человеком в глобальном масштабе достиг своего максимального предела - 100 млн т в год - и не может быть существенно увеличен.
Если вылов рыбы увеличить нельзя, то напрашивается, казалось бы, очевидное решение: необходимо наращивать искусственное воспроизводство рыб и водных беспозвоночных - аквакультуру. Действительно, во многих странах мира продукция аквакультуры увеличивается быстрыми темпами и, по некоторым оценкам, уже составляет почти половину от мировых уловов: около 50 млн т (Алимов и др., 2008).
При всей важности и необходимости подъёма продукции аквакультуры имеются два потенциальных ограничения её чрезмерного роста. Первое ограничение связано с негативным воздействием аквакультуры на природные водные экосистемы. Понятно, что для аквакультуры используются участки морей, озёр и рек, в которых также нагуливается и добывается «дикая» рыба, всё ещё составляющая основу нашего рациона. Аквакультура создаёт мощное органическое и биологическое загрязнение водоёмов, последствия которого для природных экосистем ещё до конца не выявлены. Хотя существует прудовое рыбоводство, не наносящее ущерба естественным водоёмам. Во-вторых, рыбе, как и большинству других животных, для роста и развития необходимы длинноцепочечные ПНЖК, в первую очередь ДГК, при недостатке которых не развивается мозг, глаза, малёк становится неспособным к питанию и т.д. Поэтому важнейшее условие для высокопродуктивной аквакультуры - это наличие кормов, богатых ПНЖК. Эти корма производятся из беспозвоночных и рыб, добываемых из природных экосистем. Например, аквакультура сёмги в Норвегии потребляет больше биомассы рыб, чем производит (Pauly et al., 2002). Получается своеобразный замкнутый круг: чем больше мы хотим произвести рыбы, богатой ПНЖК, в аквакультуре, тем больше мы должны выловить богатой ПНЖК рыбы и беспозвоночных из природных экосистем. В настоящее время предпринимаются шаги для искусственного производства ЭПК и ДГК вне природных водоёмов: в промышленной культуре микроводорослей, а также путём выращивания генетически модифицированных высших растений, в которых клонированы гены синтеза длинноцепочечных омега-3 ПНЖК из некоторых микроорганизмов (водорослей и бактерий) (Robert, 2006; Damude, Kinney, 2007). Однако экономические перспективы этих мероприятий пока не ясны. Уже сейчас полноценный сбалансированный корм - один из самых затратных компонентов аквакультуры. Стоимость рыбы, выращиваемой на искусственно произведённых кормах, может оказаться не по карману массовому потребителю.
Очевидно, что даже при самом успешном развитии аквакультуры природные экосистемы останутся одним из основных источников рыбопродукции для человека. Оценкой продукционного потенциала природных водоёмов и созданием научных предпосылок для его рациональной эксплуатации занимается современная экологическая наука о водных экосистемах - гидробиология. Сам термин «продукционная гидробиология» и научные основы для количественной оценки биологической продукции водных экосистем были созданы в СССР под руководством выдающегося учёного, члена-корреспондента АН СССР Г.Г. Винберга. В настоящее время научную школу продукционной гидробиологии в России возглавляет академик РАН А.Ф. Алимов. Теория продукционной гидробиологии позволяет рассчитать величины первичной продукции, производимой микроводорослями, на этой основе вычислить продукцию водных беспозвоночных - зоопланктона и зообентоса, являющихся кормом для рыб, и затем определить рыбную продукцию, которая может быть изъята из конкретной водной экосистемы без ущерба для популяций рыб и их дальнейшего успешного воспроизводства (Алимов, 1989).
При всей важности количественных оценок возможного вылова рыбы, производимых современной продукционной гидробиологией, в современных условиях важнейшее значение приобретают знания о биохимическом качестве добываемой рыбной продукции, а именно о содержании в ней длинноцепочечных омега-3 ПНЖК. Как уже говорилось, далеко не все микроводоросли способны синтезировать ЭПК и ДГК. Таким образом, в различных водных экосистемах по трофическим цепям будет передаваться на верхние уровни - к рыбе - разное количество ЭПК и ДГК. То есть в разных водных экосистемах водится рыба разного биохимического качества. Очевидно, что представления о ценной - «красной» - рыбе и малоценных породах зародились за много веков до изобретения хроматомасс-спектрометров (рис. 8) и открытия омега-3 ПНЖК (рис. 7). Однако эти представления о ценности тех или иных видов рыб удивительным образом совпали с содержанием в них ЭПК и ДГК. О пищевой ценности конкретных рыб речь пойдёт ниже, а пока лишь заметим, что содержание ЭПК+ДГК в их мясе может различаться более чем в 40 раз (табл. 2). Очевидно, что если в аквакультуре или в уловах один вид рыбы заменится на другой, то при одной и той же биомассе количество добываемых ПНЖК может также измениться в несколько раз. Следовательно, количественную оценку потенциальной рыбной продукции, которую можно извлечь из того или иного озера, моря или реки, нужно дополнить данными о качественной составляющей этой продукции, а именно о содержании в ней длинноцепочечных омега-3 ПНЖК, играющих ключевую роль в предотвращении сердечно-сосудистых заболеваний.
Таким образом, наряду с успешно решае - мой задачей оценки общего продукционного потенциала каждой эксплуатируемой водной экосистемы, определяемого по биомассе вылавливаемой рыбы, необходимо провести инвентаризацию водоёмов по их способности производить то или иное количество длинноцепочечных омега-3 ПНЖК, чрезвычайно необходимых человечеству для предотвращения эпидемий сердечно-сосудистых и нервных заболеваний. Одна из основных целей исследований в данной области - понять, в каких экосистемах может быть достигнута максимальная продукция ПНЖК. Например, в так называемых олиготрофных (малокормных) озёрах и реках с низкой температурой и чистой прозрачной водой основными видами водорослей являются диатомеи, способные к синтезу длинноцепочечных омега-3 ПНЖК.
Таблица 2. Содержание эйкозапентаеновой и докозагексаеновой кислоты (г/кг сырой массы) в некоторых видах рыб из природных местообитаний
| Вид |
ЭПК |
ДГК |
сумма |
Источник |
|||
| 1 |
2 |
3 |
4 |
5 |
|||
| Отряд Сельдеобразные |
|||||||
| Сардина (Sardinops sagax) |
6.6 |
19.0 |
25.6 |
Huynh, Kitts, 2009 |
|||
| Сельдь (Clupea harengus) |
8.5 |
8.3 |
16.8 |
Huynh, Kitts, 2009 |
|||
| Европейская алоза (Alosa alosa) |
0.12 |
0.43 |
0.5 |
Chuang et al., 2012 |
|||
| Отряд Лососеобразные |
|||||||
| Сёмга (Salmo salar) |
6.2 |
5.8 |
12.0 |
Kitson et al., 2009 |
|||
| Горбуша (Oncorhynchus gorbuscha) |
1.7 |
3.3 |
5.0 |
Gladyshev et al., 2006 |
|||
| Кумжа (Salmo trutta)* |
0.9 |
3.5 |
4.4 |
Heissenberger et al., 2010 |
|||
| Радужная форель (Oncorhynchus mykiss)* |
0.9 |
3.1 |
4.0 |
Heissenberger et al., 2010 |
|||
| Арктический голец (Salvelinus alpinus)* |
1.3 |
2.8 |
4.1 |
Heissenberger et al., 2010 |
|||
| Американский сиг (Coregonus clupeaformis)* |
0.7 |
2.4 |
3.1 |
Wagner et al., 2010 |
|||
| Хариус европейский (Thymallus thymallus) |
0.9 |
2.0 |
2.9 |
Ahlgren et al., 1994 |
|||
| Хариус сибирский (Thymallus arcticus) |
0.7 |
1.9 |
2.6 |
Sushchik et al., 2007 |
|||
| Нерка (Oncorhynchus nerka) |
0.8 |
1.9 |
2.6 |
Гладышев и др., 2010 |
|||
| Отряд Корюшкообразные |
|||||||
| Корюшка морская (Hypomesus pretiosus) |
3.6 |
5.7 |
9.3 |
Huynh, Kitts, 2009 |
|||
| Мойва (Mallotus villosus) |
3.6 |
4.6 |
8.2 |
Huynh, Kitts, 2009 |
|||
| Отряд Скорпенообразные |
|||||||
| Канареечный окунь (Sebastes pinniger) |
3.5 |
5.4 |
7.9 |
Huynh, Kitts, 2009 |
|||
| Красная скорпена (Scorpaena scrofa) |
0.29 |
1.4 |
1.7 |
Chuang et al., 2012 |
|||
| Отряд Окунеобразные |
|||||||
| Ставрида обыкновенная (Trachurus trachurus) |
1.64 |
5.86 |
7.5 |
Chuang et al., 2012 |
|||
| Чёрная сабля-рыба (Aphanopus carbo) |
0.8 |
2.9 |
3.7 |
Maulvault et al., 2012 |
|||
| Ласкирь (Diplodus sargus) |
1.0 |
2.3 |
3.3 |
Ozyurt et al., 2005 |
|||
| Золотистый спар (Sparus aurata) |
0.9 |
2.0 |
2.9 |
Ozyurt et al., 2005 |
|||
| Лаврак (Dicentrarchus labrax) |
0.52 |
1.75 |
2.3 |
Chuang et al., 2012 |
|||
| Барабулька (Mullus barbatus) |
0.48 |
0.94 |
1.9 |
Chuang et al., 2012 |
|||
| Окунь (Perca fluviatilis) |
0.3 |
0.9 |
1.2 |
Ahlgren et al., 1994 |
|||
| Ёрш (Gymnocephalus cernuus) |
0.4 |
0.8 |
1.2 |
Ahlgren et al., 1994 |
|||
| Судак (Sander lucioperca) |
0.2 |
0.8 |
1.1 |
Ahlgren et al., 1994 |
|||
| Тиляпия (Tilapia zilli) |
0.1 |
0.5 |
0.7 |
Kwetegyeka et al., 2008 |
|||
| Нильская тиляпия (Oreochromis niloticus) |
0.1 |
0.6 |
0.7 |
Kwetegyeka et al., 2008 |
|||
| Нильский окунь (Lates niloticus) |
0.1 |
0.5 |
0.6 |
Kwetegyeka et al., 2008 |
|||
| Горбыль темный (Sciaena umbra) |
0.05 |
0.19 |
0.2 |
Chuang et al., 2012 |
|||
| Пеламида атлантическая (Sarda sarda) |
0.03 |
0.15 |
0.2 |
Chuang et al., 2012 |
|||
| Отряд Угреобразные |
|||||||
| Угорь (Anguilla anguilla) |
1.6 |
2.2 |
3.7 |
Ahlgren et al., 1994 |
|||
| Отряд Трескообразные |
|||||||
| Минтай (Theragra chalcogramma) |
1.0 |
2.4 |
3.4 |
Huynh, Kitts, 2009 |
|||
| 1 2 |
3 |
4 |
5 |
||||
| Хек (Merluccius productus) 0.9 |
1.5 |
2.4 |
Huynh, Kitts, 2009 |
||||
| Треска (Gadus morhua) 0.6 |
1.5 |
2.1 |
Gladyshev et al., 2007 |
||||
| Налим (Lota lota) 0.5 |
0.9 |
1.3 |
Ahlgren et al., 1994 |
||||
| Мерлан (Gadus merlangus) 0.08 |
0.48 |
0.6 |
Chuang et al., 2012 |
||||
| Отряд Камбалообразные |
|||||||
| Камбала (Lepidopsetta bilineata) 1.8 |
1.1 |
2.9 |
Gladyshev et al., 2007 |
||||
| Отряд Карпообразные |
|||||||
| Язь (Leuciscus idus) 0.5 |
1.1 |
1.6 |
Ahlgren et al., 1994 |
||||
| Карась серебряный (Carassius gibelio) 0.6 |
1.0 |
1.6 |
Rogozin et al., 2011 |
||||
| Плотва (Rutilus rutilus) 0.4 |
1.0 |
1.4 |
Ahlgren et al., 1994 |
||||
| Густера (Blicca bjoerkna) 0.4 |
0.8 |
1.2 |
Ahlgren et al., 1994 |
||||
| Лещ (Abramis brama) 0.4 |
0.6 |
1.0 |
Ahlgren et al., 1994 |
||||
| Линь (Tinca tinca) 0.3 |
0.5 |
0.8 |
Ahlgren et al., 1994 |
||||
| Карась золотой (Carassius carassius) 0.2 |
0.6 |
0.8 |
Ahlgren et al., 1994 |
||||
| Отряд Щукообразные |
|||||||
| Щука (Esox lucius) 0.3 |
1.0 |
1.3 |
Ahlgren et al., 1994 |
||||
| Отряд Сомообразные |
|||||||
| Викторианский сомик (Synodontis victoriae) 0.2 |
0.7 |
0.9 |
Kwetegyeka et al., 2008 |
||||
| Суданский сом (Bagrus docmas) 0.1 |
0.7 |
0.8 |
Kwetegyeka et al., 2008 |
||||
| Длинноусый сом (Clarias gariepinus) 0.2 |
0.5 |
0.7 |
Kwetegyeka et al., 2008 |
||||
| Отряд Кефалеобразные |
|||||||
| Кефаль-лобан (Mugil cephalus) 0.46 |
0.08 |
0.5 |
Chuang et al., 2012 |
||||
| Отряд Двулёгочникообразные |
|||||||
| Большой протоптер (Protopterus aethiopicus) 0.1 |
0.3 |
0.4 |
Kwetegyeka et al., 2008 |
||||
| Отряд Сарганообразные |
|||||||
| Сарган (Belone belone) 0.01 |
0.15 |
0.2 |
Chuang et al., 2012 |
||||
*Пересчитано из сухого веса с использованием влажности для лососеобразных 72.5 % (Gladyshev et al., 2006, 2007; Sushchik et al., 2007).
Однако в чистой воде этих озёр содержится очень мало элементов минерального питания и продукция фотосинтеза водорослей в целом невелика. В олиготрофных озёрах и реках водятся лососеобразные рыбы с очень высоким содержанием ПНЖК (табл. 2), но их уловы являются сравнительно низкими. В то же время в теплых мутных озёрах, называемых эвтрофными, преимущественно развиваются зелёные и сине-зелёные водоросли, не способные к синтезу ЭПК и ДГК. Они получают достаточное количество минеральных элементов и дают очень высокую первичную продукцию. На этой продукции вырастают карпообразные рыбы, для которых характерно низкое содержание ПНЖК (табл. 2), но их уловы весьма высоки. Понятно, что количество ЭПК+ДГК, получаемое человеком из олиготрофных и эвтрофных озёр, может быть в конечном итоге одина ково низким. Между этими двумя крайними вариантами трофических типов озёр имеются промежуточные варианты: мезотрофные водоёмы (рис. 13). Предстоит изучить все эти варианты и выяснить, какой из них наиболее выгоден для человека с точки зрения получе - ния максимальной продукции незаменимых длинноцепочечных омега-3 ПНЖК. Подобная научная задача ставится нами впервые в мировой практике. Всем хорошо известно, что существует Красная книга, содержащая перечень особо охраняемых видов животных и растений. Следует создать подобный перечень («красную книгу») водных экосистем, являющихся наиболее важными источниками необходимых человеку ПНЖК и подлежащих особой охране. Очевидно, что подобный перечень должен опираться на надёжные данные широкомасштабных научных исследований.
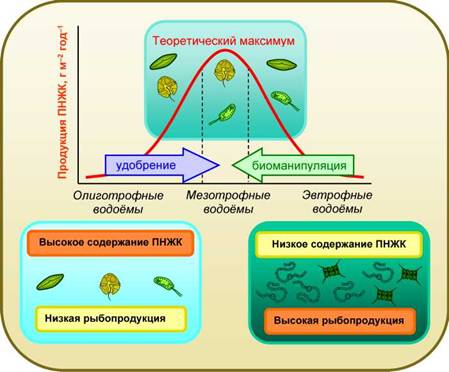
Рис. 13. Гипотетическая зависимость продукции ПНЖК от трофического статуса водных экосистем
Как можно на практике увеличить продукцию ПНЖК в водных экосистемах? В мо - рях это достигается исключительно за счёт оптимизации вылова и охраны морских вод от загрязнения. Во внутренних водоёмах имеются дополнительные возможности управления рыбной продукцией. Олиготрофные водоёмы можно удобрять, добавляя в них элементы минерального питания - азот и фосфор - в строго определённых количествах. Подобные эксперименты проводятся, например, в Канаде: в последнее десятилетие минеральные удобрения вносятся в олиготрофное оз. Кутенай (площадь 395 км2) для увеличения продукции популяций лососевых рыб, таких как нерка (кокани), форель и голец (Hyatt et al., . Успешный эксперимент по удобрению проведён недавно и на субальпийском олиго- трофном водохранилище Стора Мелкваттнет в Швеции (Persson et al., 2008).

Рис. 14. Летнее «цветение» воды сине-зелёными водорослями (цианобактериями) в эвтрофном водохранилище Бугач, расположенном в окрестностях г. Красноярска
Эвтрофные водоёмы, для которых характерно «цветение» воды сине-зелёными водорослями (цианобактериями) (рис. 14), могут быть очищены от этих вредоносных микроорганизмов путём так называемой биоманипуляции «top-down» (Гладышев, 2001). Суть биоманипуляции состоит в целенаправленном изменении трофической структуры водоёма, а именно в увеличении численности хищных рыб и уменьшении численности рыб, питающихся планктонными и бентосными беспозвоночными (рис. 15). Численность мелких беспозвоночных при этом увеличивается, и они поедают «лишние» микроводоросли, тем самым останавливая их «цветение» (рис. 15). Впервые в России биоманипуляция «top-down» была подготовлена и осуществлена в 2002-2003 годах на небольшом рекреационном водохранилище (Гладышев и др., 2006). В результате интродукции в водохранилище щуки и двукратного снижения численности карася прекратилось «цветение» сине-зелёных водорослей Microcystis (рис. 14) и начался рост перидиниевых водорослей, способных к синтезу ЭПК и ДГК. Понятно, что снижение биомассы сине-зелёных водорослей в результате биоманипуляции является лишь первым шагом к повышению ценной рыбопродукции. Здесь необходимы дальнейшие полномасштабные гидробиологические, ихтиологические и биохимические исследования.
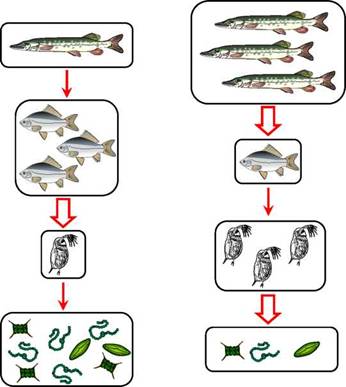
Рис. 15. Схема биоманипуляции трофическими цепями в водных экосистемах для уменьшения биомассы фитопланктона. Увеличение численности хищника (справа) приводит к уменьшению биомассы фитопланктона (эффект трофического каскада)
Таким образом, в настоящее время имеются потенциальные возможности для повышения продукции ПНЖК в озёрах и водохранилищах путём перевода олиготрофных и эвтрофных водоёмов в статус мезотрофных, в которых предположительно может наблюдаться максимум продукции ЭПК и ДГК. Однако любое вмешательство в природу требует предварительного проведения тщательных научных исследований. Возможным вариантом повышения качества рыбопродукции является зарыбление ценными видами. Например, в озёра юга Хакасии многие годы зарыбляется пелядь, которая относится к лососеобразным рыбам и предположительно имеет высокое содержание ЭПК и ДГК. Пелядь в этих озёрах, ранее бывших практически безрыбными, быстро достигает товарных размеров и хорошо продаётся на местных рынках. Перспективным вариантом увеличения продукции ПНЖК представляется зарыбление водоёмов растительноядными рыбами, потребляющими водоросли и высшие водные растения, то есть образующими короткую пищевую цепь.
Наряду с задачами инвентаризации продукции ПНЖК в водных экосистемах и поиска научно обоснованных способов её повышения остро стоит проблема охраны водоёмов от антропогенного воздействия, приводящего к снижению производства в них полезных для человека биологически активных веществ. В настоящее время риски снижения продукции ПНЖК в природных водоёмах связаны в основном с тремя факторами: глобальным потеплением климата, антропогенным эвтро- фированием и загрязнением (рис. 16).
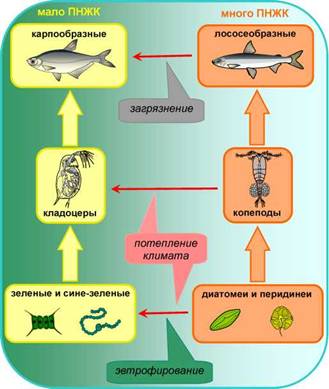
Рис. 16. Влияние антропогенных факторов на продукцию ПНЖК в водных экосистемах
Глобальное потепление, приводящее к по - вышению температуры воды, напрямую влияет на содержание ПНЖК в водных экосистемах. У водорослей, приспособленных к низким температурам, высокое содержание полинена- сыщенных жирных кислот, имеющих низкую температуру замерзания, обеспечивает оптимальную вязкость клеточных мембран, от которой зависит нормальное функционирование клетки. Чем ниже температура окружающей среды, тем больше должно содержаться в мембране «антифризов», а именно ПНЖК. При высоких температурах клеточная мембрана, наоборот, должна «затвердевать» и водоросли вместо ПНЖК начинают синтезировать насыщенные жирные кислоты с высокой температурой плавления (Guschina, Harwood, 2009). Поэтому, например, диатомовые водоросли, содержащие много ЭПК, доминируют в более холодных водах, тогда как в тёплых водах пре - имущество получают зелёные и сине-зелёные водоросли, вообще не синтезирующие длинноцепочечные ПНЖК. Подобная адаптация к температуре происходит и у беспозвоночных животных: виды, богатые ПНЖК, замещаются другими организмами. Например, в более тёплых озёрах снижение запасов ПНЖК в зоопланктоне происходит за счёт замены ко- пепод (веслоногих рачков), накапливающих в своей биомассе большое количество ДГК, на кладоцер (ветвистоусых рачков), содержащих очень мало ДГК (рис. 16) (Gladyshev et al., 2011a). Копеподы и кладоцеры служат основным кормом для мальков рыб. Как уже отмечалось, ДГК является необходимым компонентом для роста и развития рыб, поэтому снижение её концентрации в кормовых объектах автоматически приводит к снижению качества и количества рыбной продукции. Известно, что в связи с потеплением климата в озёрах Европы лососеобразные рыбы заменяются окунеобразными, а те, в свою очередь, замещаются карпообразными (Jeppesen et al., . Как видно из табл. 2, в ряду «лососевые ^ окуневые ^ карповые» происходит существенное снижение содержания ПНЖК в мясе рыб (рис. 16). Следует отметить, что в морях и океанах основу биомассы зоопланктона со - ставляют именно копеподы, богатые ПНЖК, поэтому морские пелагические рыбы (сельдь, сардина, мойва), т.е. обитающие в толще воды, где пищевая цепь начинается с планктона, как правило, имеют высокое содержание ЭПК и ДГК (табл. 2).
Наряду с потеплением климата антропогенное эвтрофирование, т.е. поступление в водоёмы минеральных элементов (азота и фосфора), также является причиной процветания сине-зелёных водорослей и подавления диатомей, богатых ПНЖК (рис. 16). Кроме азота и фосфора человек сбрасывает в водоёмы органические вещества и тяжёлые металлы. Влияние тех или иных органических загрязнений и тяжёлых металлов на продукцию ПНЖК в водных экосистемах ещё предстоит исследовать. Однако уже установлено, что, например, в р. Енисей антропогенное загрязнение приводит к снижению эффективности переноса ПНЖК по трофической цепи, т.е. продукция водорослей остаётся на прежнем уровне или даже возрастает, а вот содержание ЭПК и ДГК в кормовых организмах достоверно уменьшается (Гладышев и др., 2012). Кроме того, тяжёлые металлы, сбрасываемые в водоёмы со сточными водами, накапливаются в верхних звеньях трофической цепи, т. е. в рыбах. В этой связи возникает очень серьёзная проблема: потребляя рыбу, чтобы получить дозу необходимых для здоровья ПНЖК, человек может одновременно получить и токсичную дозу тяжёлых металлов (радионуклидов, пестицидов). Очевидно, что необходим постоянный мониторинг содержания не только полезных, но и токсических веществ в рыбной продукции. Такой мониторинг был начат для рыбы р. Енисей. Была выведена специальная формула для количественного расчёта соотношения «риск-польза» при потреблении человеком той или иной продукции. Формула позволяет рассчитать, опираясь на данные мониторинга, какое количество той или иной рыбы, вылавливаемой в конкретном водоёме, человек может употребить в пищу для профилактики сердечно-сосудистых заболеваний без риска получить при этом токсичную дозу тяжёлых металлов или иных опасных веществ (Gladyshev et al., 2009b).
Оптимальные порции рыбы и её кулинарная обработка
Как убедительно доказано современной медицинской наукой, употребление в пищу рыбы, содержащей достаточное количество ЭПК и ДГК, является для большинства людей необходимым условием нормального функционирования кровеносной и нервной систем. Содержание ПНЖК во многих видах рыб хорошо известно (табл. 2). Следует отметить, что наибольшей пищевой ценностью в отношении содержания ЭПК и ДГК обладают морские пелагические рыбы, т.е. те, которые водятся в толще поверхностных вод и их пищевые цепи основаны на продукции диатомовых и перидиниевых водорослей, синтезирующих большое количество длинноцепочечных омега-3 ПНЖК, а также на планктонных копеподах, способных накапливать в своей биомассе ПНЖК микроводорослей. Такими рыбами являются сельдь, сардина, мойва, питающиеся зоопланктоном (копеподами), и крупные лососевые (сёмга, горбуша, нерка), питающиеся мелкой пелагической рыбой. Морские придонные рыбы (например, камбала), как и почти все пресноводные рыбы, содержат относительно мало ПНЖК. Некоторые исследователи полагают, что пресноводная рыба вообще не может служить значимым источником ЭПК и ДГК для питания человека (Philibert et al., . Действительно, рыба, вылавливаемая в африканских озёрах, таких как Виктория, а именно тиляпия, нильский окунь, викторианский сомик, содержит чрезвычайно мало ПНЖК (табл. 2), и её потребление в пищу не может обеспечить рекомендованные суточные дозы ЭПК и ДГК (Kwetegyeka et al.,
Тем не менее представления о малой питательной ценности всех пресноводных рыб (Philibert et al., 2006) во многом основаны на недостаточной изученности содержания ПНЖК в видах, населяющих континентальные водоемы. В большинстве исследований до сих пор применяется косвенный метод оценки содержания ПНЖК: их количество выражается в процентах от общей суммы жирных кислот, а концентрации в единице массы продукта остаются неизвестными. Для того чтобы оценить содержание ЭПК + ДГК не в относительных (проценты), а в абсолютных единицах (мг ПНЖК / г массы рыбы), необходимо при хроматографических анализах добавлять так называемый внутренний стандарт. Однако внутренний стандарт использует лишь относительно небольшое число лабораторий. Более того, чтобы при анализах достоверно отличить ЭПК и особенно ДГК от других кислот, не представляющих уникальной физиолого-биохимической ценности, следует использовать масс-спектрометрический детектор. Но на практике данный детектор применяют весьма редко. Поэтому для уточнения данных о пищевой ценности пресноводных рыб необходимо провести широкомасштабные исследования с использованием внутреннего стандарта и масс-спектрометрии. Можно с уверенностью предположить, что сиговые рыбы, относящиеся к отряду Лососеобразных и питающиеся планктонными копеподами, такие как омуль, пелядь, ряпушка, будут иметь весьма высокое содержание ЭПК и ДГК.
Однако все обсуждаемые выше данные относятся к сырой рыбе. В большинстве стран сырая рыба употребляется в пищу достаточно редко. Возникает весьма важный вопрос: как та или иная кулинарная обработка рыбы влияет на содержание в ней ЭПК и ДГК? Известно, что полиненасыщенные жирные кислоты, имеющие несколько двойных связей, являются неустойчивыми, например, к окислению или нагреванию. Поэтому долгое время считалось, что при кулинарной обработке двойные связи в ПНЖК «рвутся» и они теряют свои уникальные полезные свойства. Действительно, если нагреть химически чистую ПНЖК, она деградирует. Однако достаточно недавно с применением внутреннего стандарта и хромато-масс-спектрометрии было установлено, что в процессе кулинарной обработки в жареной, варёной и запеченной рыбе содержание ЭПК и ДГК не уменьшается по сравнению с рыбой сырой (Gladyshev et al., 2006, 2007). В рыбе ЭПК и ДГК содержатся не в чистом виде, а в составе фосфолипидов клеточных мембран (рис. 5). То есть они плотно упакованы в бинарные слои и окружены белками. Очевидно, такая «упаковка» предотвращает деградацию длинноцепочечных омега-3 ПНЖК при кулинарной обработке. Ещё более удивительным оказался тот факт, что даже в консервах содержание ЭПК и ДГК в рыбе не только не снижается, но, наоборот, увеличивается (Gladyshev et al., 2009c). Увеличение происходит потому, что при консервировании из рыбы уходит часть воды и лёгких короткоцепочечных кислот, а ПНЖК как были, так и остаются в клеточных мембранах. Таким образом, человеку следует употреблять в пищу мясо рыбы, а не вытопленный из неё жир. Кстати сказать, тот жир, который вытекает из рыбы, почти не содержит ЭПК и ДГК, поскольку он представляет собой запасные питательные липиды - триглицерины, а ПНЖК, как мы уже точно знаем, находятся в составе структурных липидов - фосфолипидов, формирующих мембраны клеток.
По результатам исследований содержания ПНЖК в готовых продуктах может быть составлена таблица, в которой указано, какую порцию того или иного блюда необходимо употребить, чтобы получить суточную дозу
Таблица 3. Порция продукта, которую необходимо употребить в пищу, чтобы получить рекомендованную суточную дозу ЭПК+ДГК, равную 1 г
Продукт Порция (г) Источник
| Сайра консервированная |
41 |
Gladyshev et al., 2009с |
| Семга жареная |
51 |
Sioen et al., 2006 |
| Сельдь консервированная |
56 |
Gladyshev et al., 2009с |
| Горбуша варёная |
167 |
Gladyshev et al., 2006 |
| Форель варёная |
175 |
Gladyshev et al., 2007 |
| Горбуша припущенная |
189 |
Gladyshev et al., 2006 |
| Горбуша запечёная |
200 |
Gladyshev et al., 2006 |
| Горбуша жареная |
233 |
Gladyshev et al., 2006 |
| Форель жареная |
244 |
Gladyshev et al., 2007 |
| Треска жареная |
246 |
Sioen et al., 2006 |
| Сельдь варёная |
256 |
Gladyshev et al., 2007 |
| Сельдь жареная |
263 |
Gladyshev et al., 2007 |
| Камбала варёная |
278 |
Gladyshev et al., 2007 |
| Камбала жареная |
323 |
Gladyshev et al., 2007 |
| Треска варёная |
417 |
Gladyshev et al., 2007 |
| Креветки жареные |
563 |
Simon et al., 2012 |
| Свинина жареная |
3333 |
Haak et al., 2007 |
ЭПК и ДГК в 1 г, рекомендованную Всемирной организацией здравоохранения и многими национальными медицинскими организациями для профилактики сердечно-сосудистых и нервных заболеваний (табл. 3). Как видно из таблицы, лидером по содержанию ПНЖК является консервированная сайра: чтобы получить 1 г ЭПК+ДГК, человеку достаточно съесть всего около 40 г этого продукта. Важно отметить, что большинство видов рыб, присутствующих на наших прилавках, при традиционных способах приготовления являются ценным источником физиологически значимых длинноцепочечных омега-3 ПНЖК (табл. 3).
Заключение
Итак, генотип большинства людей изначально запрограммирован на соотношение n-6:n-3 ПНЖК в пище около 1:1. Данное соотношение обеспечивает необходимый баланс в организме человека эндогормонов - эйкозано- идов (простагландинов, тромбоксанов, лейко- триенов), обеспечивающих нормальное функционирование сердечно-сосудистой системы, ряда других систем, органов и тканей. Однако в современных условиях индустриализации сельского хозяйства в рационе человека преобладают продукты, богатые n-6 ПНЖК. Рост соотношения n-6:n-3 в рационе населения экономически развитых стран до 20:1 сопровождался резким увеличением сердечнососудистых заболеваний, а также нервных расстройств. Для профилактики этих заболеваний необходимо ежедневно употреблять в пищу около 1 г ЭПК+ДГК. Основным источником ЭПК и ДГК на нашей планете являются водные экосистемы. Человечество должно озаботиться сохранением водных экосистем и увеличением их полезной продукции.
Н.Н. Сущик за ценные замечания и ре- стерства образования и науки РФ.
Список литературы
Алимов А.Ф. (1989) Введение в продукционную гидробиологию. Л.: Гидрометеоиздат, 152 с.
Алимов А.Ф., Алексеев А.П., Бергер В.Я. (2008) Марикультура как способ увеличения промысловых ресурсов Белого моря. Вестник РАН 78: 792-799.
Гладышев М.И. (2001) Биоманипуляция как инструмент управления качеством воды в континентальных водоемах (обзор литературы 1990-1999 гг.). Биология внутренних вод 2: 3-15.
Гладышев М.И., Анищенко О.В., Сущик Н.Н., Калачёва Г.С., Грибовская И.В., Агеев А.В. (2012) Влияние антропогенного загрязнения на содержание незаменимых полиненасы- щенных жирных кислот в звеньях трофической цепи речной экосистемы. Сибирский экологический журнал 4: 511-521.
Гладышев М.И., Лепская Е.В., Сущик Н.Н., Махутова О.Н., Калачёва ГС. (2010) Различия жирнокислотного состава покатной молоди и вернувшихся из моря взрослых особей нерки. Доклады АН 430: 548-551.
Гладышев М.И., Чупров С.М., Колмаков В.И., Дубовская О.П., Кравчук Е.С., Иванова Е.А., Трусова М.Ю., Сущик Н.Н., Калачева Г.С., Губанов В.Г., Прокопкин И.Г., Зуев И.В., Махутова О.Н. (2006) Биоманипуляция “top-down” в небольшом сибирском водохранилище без дафний. Сибирский экологический журнал 13: 31-41.
Попов Н.П. (2012) Вымирает ли Россия? Вестник РАН 82: 3-18.
Ahlgren G., Blomqvist P., Boberg M, Gustafsson I.-B. (1994) Fatty acid content of the dorsal muscle - an indicator of fat quality in freshwater fish. Journal of Fish Biology 45: 131-157.
Batista C., Barros L., Carvalho A.M., Ferreira I.C.F.R. (2011) Nutritional and nutraceutical potential of rape (Brassica napus L. var. napus) and ‘’tronchuda’’ cabbage (Brassica oleraceae L. var. costata) inflorescences. Food and Chemical Toxicology 49: 1208-1214.
Bazan N.G. (2009) Cellular and molecular events mediated by docosahexaenoic acid-derived neuroprotectin D1 signaling in photoreceptor cell survival and brain protection. Prostaglandins, Leukotrienes and Essential Fatty Acids 81: 205-211.
Bell M.V., Tocher D.R. (2009) Biosynthesis of polyunsaturated fatty acids in aquatic ecosystems: general pathways and new directions. In: Lipids in aquatic ecosystems. Arts M.T., Kainz M., Brett M.T., Eds. New York: Springer, p. 211-236.
Broadhurst C.L., Wang Y., Crawford M.A., Cunnane S.C., Parkington J.E., Schmidt W.F. (2002) Brain-specific lipids from marine, lacustrine, or terrestrial food resources: potential impact on early African Homo sapiens. Comparative Biochemistry and Physiology B 131: 653-673.
Cohen Z., Norman H.A., Heimer Y.M. (1995) Microalgae as a source of <a-3 fatty acids. In: Plants in human nutrition. Simopoulos A.P., Ed. World review of nutrition and dietetics, V. 77, Basel: Karger, p. 1-31.
Crawford M.A., Bloom M., Broadhurst C.L., Schmidt W.F., Cunnane S.C., Galli C., Gehbremeskel K., Linseisen F., Lloyd-Smith J., Parkington J. (1999) Evidence for the unique function of docosahexaenoic acid during the evolution of the modern hominid brain. Lipids 34: S39-S47.
Chuang L.T., Bulbul U., Wen P.C., Glew R.H., Ayaz F.A. (2012) Fatty acid composition of 12 fish species from the Black Sea. Journal of Food Science 77: C512-C518.
Damude H.G., Kinney A.J. (2007) Engineering oilseed plants for a sustainable, land-based source of long chain polyunsaturated fatty acids. Lipids 42: 179-185.
Davis B.C, Kris-Etherton P.M. (2003) Achieving optimal essential fatty acid status in vegetarians: current knowledge and practical implications. American Journal of Clinical Nutrition 78(suppl): 640S-6S.
Gladyshev M.I., Sushchik N.N., Gubanenko G.A., Demirchieva S.M., Kalachova, G.S. (2006) Effect of way of cooking on content of essential polyunsaturated fatty acids in muscle tissue of humpback salmon (Oncorhynchus gorbuscha). Food Chemistry 96: 446-451.
Gladyshev M.I., Sushchik N.N., Gubanenko G.A., Demirchieva S.M., Kalachova G.S. (2007) Effect of boiling and frying on the content of essential polyunsaturated fatty acids in muscle tissue of four fish species. Food Chemistry 101: 1694-1700.
Gladyshev M.I., Arts M.T., Sushchik N.N. (2009a) Preliminary estimates of the export of omega-3 highly unsaturated fatty acids (EPA+DHA) from aquatic to terrestrial ecosystems. In: Lipids in aquatic ecosystems. Arts M.T., Kainz M., Brett M.T., Eds. New York: Springer, p. 179-209.
Gladyshev M.I., Sushchik N.N., Anishchenko O.V., Makhutova O.N., Kalachova G.S., Gribovskaya I.V. (2009b) Benefit-risk ratio of food fish intake as the source of essential fatty acids vs. heavy metals: A case study of Siberian grayling from the Yenisei River. Food Chemistry 115: 545-550.
Gladyshev M.I., Sushchik N.N., Makhutova O.N., Kalachova G.S. (2009c) Content of essential polyunsaturated fatty acids in three canned fish species. International Journal of Food Science and Nutrition 60: 224-230.
Gladyshev M.I., Semenchenko V.P., Dubovskaya O.P., Fefilova E.B., Makhutova O.N., Buseva Z.F., Sushchik N.N., Razlutskij V.I., Lepskaya E.V., Baturina M.A., Kalachova G.S., Kononova O.N. (2011а) Effect of temperature on contents of essential highly unsaturated fatty acids in freshwater zooplankton. Limnologica 41: 339-347.
Gladyshev M.I., Sushchik N.N., Anishchenko O.V., Makhutova O.N., Kolmakov V.I., Kalachova G.S., Kolmakova A.A., Dubovskaya O.P. (2011b) Efficiency of transfer of essential polyunsaturated fatty acids versus organic carbon from producers to consumers in a eutrophic reservoir. Oecologia 165: 521-531.
Gibbons A. (2002) Humans’ head start: new views of brain evolution. Science 296: 835-837.
Guschina I.A., Harwood J.L. (2009) Algal lipids and effect of the environment on their biochemistry. In: Lipids in aquatic ecosystems. Arts M.T., Kainz M., Brett M.T., Eds. New York: Springer, p.1-24.
Haak L., Sioen I., Raes K., Van Camp J., De Smet S. (2007) Effect of pan-frying in different culinary fats on the fatty acid profile of pork. Food Chemistry 102: 857-864.
Harris W.S., Mozaffarian D., Lefevre M., Toner C.D., Colombo J., Cunnane S.C., Holden J.M., Klurfeld D.M., Morris M.C., Whelan J. (2009) Towards establishing dietary reference intakes for eicosapentaenoic and docosahexaenoic acids. Journal of Nutrition 139: 804S-819S.
Harwood J.L. (1996) Recent advances in the biosynthesis of plant fatty acids. Biochimica et Biophysica Acta 1301: 7-56.
Heinz E. (1993) Biosynthesis of polyunsaturated fatty acids. In: Lipid metabolism in plants. Moore T.S. Ed. Boca Raton: CRC, p. 34-89.
Heissenberger M., Watzke J., Kainz M.J. (2010) Effect of nutrition on fatty acid profiles of riverine, lacustrine, and aquaculture-raised salmonids of pre-alpine habitats. Hydrobiologia 650: 243-254.
Hibbeln J.R., Nieminen L.R.G., Blasbalg T.L., Riggs J.A., Lands W.E.M. (2006) Healthy intakes of n-3 and n-6 fatty acids: estimations considering worldwide diversity. American Journal of Clinical Nutrition 83(suppl):1483S-1493S.
Huynh, M.D., Kitts D.D. (2009) Evaluating nutritional quality of pacific fish species from fatty acid signatures. Food Chemistry 114: 912-918.
Hyatt K.D., McQueen D.J., Shortreed K.S., Rankin D.P. (2004) Sockeye salmon (Oncorhynchus nerka) nursery lake fertilization: Review and summary of results. Environmental Reviews 12: 133-162.
Jeppesen E., Meerhoff M., Holmgren K., Gonzalez-Bergonzoni I., Teixeira-de Mello F., Declerck
S.A.J., De Meester L., Sondergaard M., Lauridsen T.L., Bjerring R., Conde-Porcuna J.M., Mazzeo N., Iglesias C., Reizenstein M., Malmquist H.J., Liu Z., Balayla D., Lazzaro X. (2010) Impacts of climate warming on lake fish community structure and potential effects on ecosystem function. Hydrobiologia 646:73-90.
Kitson A.P., Patterson A.C., Izadi H., Stark K.D. (2009) Pan-frying salmon in an eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) enriched margarine prevents EPA and DHA loss. Food Chemistry 114: 927-932.
Kris-Etherton P.M., Grieger J.A., Etherton T.D. (2009) Dietary reference intakes for DHA and EPA. Prostaglandins, Leukotrienes and Essential Fatty Acids 81: 99-104.
Kris-Etherton P.M., Harris W.S., Appel L.J. (2002) Fish consumption, fish oil, omega-3 fatty acids, and cardiovascular disease. Circulation 106: 2747-2757.
Kwetegyeka J., Mpango G., Grahl-Nielsen O. (2008) Variation in fatty acid composition in Muscle and heart tissues among species and populations of tropical fish in lakes Victoria and Kyoga. Lipids 43:1017-1029.
Lands W.E.M. (2009) Human life: caught in the food web. In: Lipids in aquatic ecosystems. Arts M.T., Kainz M., Brett M.T., Eds. New York: Springer, p. 327-354.
Lauritzen L., Hansen H.S., Jorgensen M.H., Michaelsen K.F. (2001) The essentiality of long chain n-3 fatty acids in relation to development and function of the brain and retina. Progress in Lipid Research 40: 1-94.
Lehninger A.L., Nelson D.L., Cox M.M. (1993) Principles ofbiochemistry. N.Y.: Worth Publishers, 1010 p.
Maulvault A.L., Anacleto P., Machado R., Amaral A., Carvalho M.L., Lourenco H.M., Nunesa M.L., Marques A. (2012) Effect of sex, maturation stage and cooking methods on the nutritional quality and safety of black scabbard fish (Aphanopus carbo Lowe, 1839). Journal of the Science of Food and Agriculture 92: 1545-1553.
McNamara R.K., Carlson S.E. (2006) Role of omega-3 fatty acids in brain development and function: Potential implications for the pathogenesis and prevention of psychopathology. Prostaglandins, Leukotrienes and Essential Fatty Acids 75: 329-349.
Muskiet F.A.J., Fokkema M.R., Schaafsma A., Boersma E.R., Crawford M.A. (2004) Is docosahexaenoic acid (DHA) essential? Lessons from DHA status regulation, our ancient diet, epidemiology and randomized controlled trials. Journal of Nutrition 134: 183-186.
Ozyurt G., Pola A., Ozkutuk S. (2005) Seasonal changes in the fatty acids of gilthead sea bream (Sparus aurata) and white sea bream (Diplodus sargus) captured in Iskenderun Bay, eastern Mediterranean coast of Turkey. European Food Research and Technology 220: 120-124.
Pauly D., Christensen V., Guenette S., Pitcher T.J., Sumaila U.R., Walters C.J., Watson R., Zeller D. (2002) Towards sustainability in world fisheries. Nature 418:689-695.
Persson J., Vrede T., Holmgren S. (2008) Responses in zooplankton populations to food quality and quantity changes after whole lake nutrient enrichment of an oligotrophic sub-alpine reservoir. Aquatic Sciences 70: 142 - 155.
Phang M., Lazarus S., Wood L.G., Garg M. (2011) Diet and thrombosis risk: nutrients for prevention of thrombotic disease. Seminars in Thrombosis and Hemostasis 37: 199-208.
Philibert A., Vanier C., Abdelouahab N., Chan H.M., Donna Mergler D. (2006) Fish intake and serum fatty acid profiles from freshwater fish. American Journal of Clinical Nutrition 84: 1299307.
Plourde M., Cunnane S.C. (2007) Extremely limited synthesis of long chain polyunsaturates in adults: implications for their dietary essentiality and use as supplements. Applied Physiology, Nutrition and Metabolism 32: 619-634.
Reis L.C., Hibbeln J.R. (2006) Cultural symbolism of fish and the psychotropic properties of omega-3 fatty acids. Prostaglandins, Leukotrienes and Essential Fatty Acids 75: 227-236.
Robert S.S. (2006) Production of eicosapentaenoic and docosahexaenoic acid-containing oils in transgenic land plants for human and aquaculture nutrition. Marine Biotechnology 8: 103 - 109.
Rodriguez-Carpena J.G., Morcuende D., Estevez M. (2012) Avocado, sunflower and olive oils as replacers of pork back-fat in burger patties: Effect on lipid composition, oxidative stability and quality traits. Meat Science 90: 106-115.
Rogozin D.Y., Pulyayevskaya M.V., Zuev I.V., Makhutova O.N., Degermendzhi A.G. (2011) Growth, diet and fatty acid composition of Gibel carp Carassius gibelio in Lake Shira, a brackish water body in Southern Siberia. Journal of Siberian Federal University. Biology 4: 86-103.
Roth G., Dicke U. (2005) Evolution of the brain and intelligence. Trends in Cognitive Sciences 9: 250-257.
Rule D.C., Broughton K.S., Shellito S.M., Maiorano G. (2002) Comparison of muscle fatty acid profiles and cholesterol concentrations of bison, beef cattle, elk, and chicken. Journal of Animal Sciences 80: 1202-1211.
Saldanha L.G., Salem Jr. N., Brenna J.T. (2009) Workshop on DHA as a required nutrient: Overview. Prostaglandins, Leukotrienes and Essential Fatty Acids 81: 233-236.
Sanders T.A.B. (2000) Polyunsaturated fatty acids in the food chain in Europe. American Journal of Clinical Nutrition 71(suppl): 176S-8S.
SanGiovanni J.P., Chew E.Y. (2005) The role of omega-3 long-chain polyunsaturated fatty acids in health and disease of the retina. Progress in Retinal and Eye Research 24: 87-138.
Simopoulos A.P. (2000) Human requirement for n-3 polyunsaturated fatty acids. Poultry Science 79: 961-970.
Simon S.J.G.B., Sancho R.A.S., Lima F.A., Cabral C.C.V.Q., Souza T.M., Bragagnolo N., Lira G.M. (2012) Interaction between soybean oil and the lipid fraction of fried pitu prawn. LWT - Food Science and Technology 48: 120-126.
Sinclair A.J., Attar-Bashi N.M., Li D. (2002) What is the role of a-linolenic acid for mammals? Lipids 37: 1113-1123.
Sioen I., Haak L., Raes K., Hermans C., De Henauw S., De Smet S., Van Camp J. (2006) Effects of pan-frying in margarine and olive oil on the fatty acid composition of cod and salmon. Food Chemistry 98: 609-617.
Stark A.H., Crawford M.A., Reifen R. (2008) Update on alpha-linolenic acid. Nutrition Reviews 66: 326-332.
Suchowilska E., Wiwart M., Borejszo Z., Packa D., Kandler W., Krska R. (2009) Discriminant analysis of selected yield components and fatty acid composition of chosen Triticum monococcum, Triticum dicoccum and Triticum spelta accessions. Journal of Cereal Science 49: 310-315.
Sushchik N.N., Gladyshev M.I., Kalachova G.S. (2007) Seasonal dynamics of fatty acid content of a common food fish from the Yenisei River, Siberian grayling, Thymallus arcticus. Food Chemistry 104: 1353-1358.
Tocher D.R., Leaver M.J., Hodson P.A. (1998) Recent advances in the biochemistry and molecular biology of fatty acyl desaturase. Progress in Lipid Research 37: 73-117.
Usydus Z., Szlinder-Richert J., Adamczyk M., Szatkowska U. (2011) Marine and farmed fish in the Polish market: Comparison of the nutritional value. Food Chemistry 126: 78-84.
Wagner T., Jones M.L., Ebener M.P., Arts M.T., Brenden T.O., Honeyfield D.C., Wright G.M., Faisal M. (2010) Spatial and temporal dynamics of lake whitefish (Coregonus clupeaformis) health indicators: Linking individual-based indicators to a management-relevant endpoint. Journal of Great Lakes Research 36: 121-134.
Wall R., Ross R.P., Fitzgerald G.F., Stanton C. (2010) Fatty acids from fish: the anti-inflammatory potential of long-chain omega-3 fatty acids. Nutrition Reviews 68: 280-289.
Wood J.D., Enser M., Fisher A.V., Nute G.R., Sheard P.R., Richardson R.I., Hughes S.I., Whittington
M. (2008) Fat deposition, fatty acid composition and meat quality: A review. Meat Science 78: 343-358.