| Похожие рефераты | Скачать .docx | Скачать .pdf |
Реферат: Меристемы или образовательные ткани
МЕРИСТЕМЫ ИЛИ ОБРАЗОВАТЕ ЛЬНЫЕ ТКАНИ
Цитоплазма. Химический состав, физическое состояние, строение и свойства. Поступление веществ в клетку. Типы движения цитоплазмы.
Цитоплазма бесцветная, имеет слизистую консистенцию и содержит различные вещества, в том числе и высокомолекулярные соединения, например белки, присутствие которых обусловливает коллоидные свойства цитоплазмы. Цитоплазма – часть протопласта, заключенная между плазмалеммой и ядром. Основу цитоплазмы составляет ее матрикс, или гиалоплазма, - сложная бесцветная, оптически прозрачная коллоидная система, способная к обратимым переходам из золя в гель.
В цитоплазме растительных клеток имеются органоиды: небольшие тельца, выполняющие специальные функции, - пластиды, комплекс Гольджи, эндоплазматическая сеть, митохондрии и т.д. В цитоплазме осуществляется большая часть процессов клеточного метаболизма, исключая синтез нуклеиновых кислот, происходящих в ядре. Цитоплазма пронизана мембранами – тончайшими (4-10нм) пленками, построенными в основном из фосфолипидов и липопротеинов. Мембраны ограничивают цитоплазму от клеточной оболочки и вакуоли и внутри цитоплазмы образуют эндоплазматическую сеть (ретикулум) – систему мелких вакуолей и канальцев, соединенных друг с другом.
Важнейшим свойством цитоплазмы, связанным прежде всего с физико-химическими особенностями гиалоплазмы, является ее способность к движению. В клетках с одной крупной вакуолью движение осуществляется обычно в одном направлении (циклоз) за счет особых органоидов – микрофиламентов, представляющих собой нити особого белка - актина. Движущая гиалоплазма увлекает пластиды и митохондрии. Клеточный сок, находящийся в вакуолях, представляет собой водный раствор различных веществ: белков, углеводов, пигментов, органических кислот, солей, алкалоидов и т.п. Концентрация веществ, находящихся в клеточном соке, обычно выше концентрации веществ во внешней среде (почва, водоемы). Различие концентраций в известной мере обусловливает возможность поступления в клетку воды и почвенных растворов, что до некоторой степени объясняется явлением осмоса. В клетке роль полупроницаемой мембраны играет цитоплазма. Пограничные слои цитоплазмы, выстилающие оболочку и клеточную вакуоль, проницаемы только для воды и некоторых растворов, но непроницаемы для многих растворенных в воде веществ. Это свойство цитоплазмы получило название полупроницаемости или избирательной проницаемости. В отличие от цитоплазмы клеточная оболочка проницаема для всех растворов, непроницаема она только для твердых частиц. Поступление веществ в клетку нельзя сводить только к осмотическим явлениям, которые выражены во взрослых клетках с хорошо развитыми вакуолями. В действительности это очень сложный процесс, обусловленный многими факторами. Активное участие в поглощении веществ принимает вся система коллоидов цитоплазмы. Интенсивность движения зависит от температуры, степени освещения, снабжения кислородом и т.д.
В очень молодых клетках цитоплазма заполняет почти всю их полость. По мере роста клетки в цитоплазме появляются мелкие вакуоли, заполненные клеточным соком, представляющим собой водный раствор различных органических веществ. Впоследствии, при дальнейшем росте клетки, вакуоли увеличиваются в размерах и, сливаясь, часто образуют одну большую центральную вакуоль, оттесняющую цитоплазму к оболочке клетки. В таких клетках все органоиды располагаются в тонком постенном слое цитоплазмы. Иногда ядро остается в центре клетки. В этом случае цитоплазма, образующая вокруг него ядерный кармашек, соединяется с постенным слоем тонкими цитоплазматическими тяжами.
В слое цитоплазмы расположены хлоропласты, выстилающие верхнюю стенку. Они представляют собой почти округлые или слегка овальные тельца. Изредка можно встетить пластиды, перетянутые посередине.
Понятие о тканях. Классификация тканей. Отличие постоянных тканей от образовательных.
У большинства наземных растений клетки тела неодинаковы по своим функциям, строению и происхождению. Эта разнородность возникла и закрепилась, в процессе приспособления растений к различиям воздушной и почвенной сред. Системы клеток, структурно и функционально сходны друг с другом и обычно имеющие общее происхождение, получили название ткачей. Ткани имеются почти у всех высших растений. Нет их только у части моховидных (печеночники). Водоросли и багрянки (низшие растения), как правило, также не имеют развитых тканей.
Комплексы клеток, сходных по функциям, а большей частью и по строению, имеющих одинаковое происхождение и определенную локализацию в теле растения, называют тканями.
Распределение тканей в органах растений и их структура тесно связаны с выполнением ими определенных физиологических функций.
Ткани, состоящие из одного типа клеток, получили название простых, а состоящие из разных типов клеток — сложных, или комплексных. Существуют различные классификации тканей, но все они достаточно условны. Растительные ткани делят на несколько групп в зависимости от основной функции:
1) меристемы, или образовательные ткани (ткани состоящие из живых тонкостенных, интенсивно делящихся клеток);
а) верхушечные (апикальные) меристимы (расположенны на верхушках стеблей и в окончаниях корней) обусловливают рост этих органов в длину;
б) боковые мерестимы – камбий и феллоген (камбий обеспечивает утолщение стебля и корня. Феллоген образует пробку)
2) покровные (защищают внутренние ткани растений от прямого влияния внешней среды, регулируют испарение и газообмен)
а) эпидермис;
б) пробка;
3) проводящие (обеспечивают проведение воды, почвенных растворов и продуктов ассимиляции, вырабатываемых листьями. Проводящие ткани по происхождению могут быть первичными и вторичными.);
а) кселима или древесная ткань (ткань проводящая воду)
б) флоэма или луб (ткань проводящая органические вещества, образованные растением в процессе фотосинтеза);
4) механические (обусловливают прочность растения);
а) коленхима (состоит из паренхимы или несколько удлиненных клеток с неравномерно утолщенными целлюлозными стенками);
б) склеренхима (клетки имеют равномерно утолщенные одревесневшие стенки);
1) волокна;
2) склериды;
5) основные (состоящие из однородных паренхимных клеток, которые заполняют пространство между другими тканями);
6) секреторные, или выделительные (содержащие продукты отброса).
Лишь клетки меристематических тканей способны к делению. Клетки прочих тканей, как правило, к делению неспособны, и их число увеличивается за счет деятельности соответствующих меристем. Такие ткани называют постоянными. Постоянные ткани возникают из меристем в результате клеточной дифференцировки. Дифференцировка заключается в том, что в ходе индивидуального развития организма (онтогенеза) возникают качественные различия между первоначально однородными клетками, при этом изменяются строение и функциональные свойства клеток. Обычно дифференцировка необратима. На ход ее оказывают влияние вещества, выполняющие роль гормонов.
МЕРИСТЕМЫ ИЛИ ОБРАЗОВАТЕ ЛЬНЫЕ ТКАНИ
Меристемы (от греч. «меристос» — делимый), или образовательные ткани, обладают способностью к делению и образованию новых клеток. За счет меристем формируются все прочие ткани и осуществляется длительный (в течение всей жизни) рост растения. У животных меристемы отсутствуют, чем объясняется ограниченный период их роста. Клетки меристем отличаются высокой метаболической активностью. Одни клетки меристем, получившие название инициальных, задерживаются на эмбриональной стадии развития в течение всей жизни растения, другие постепенно дифференцируются и превращаются в клетки различных постоянных тканей. Инициальная клетка меристемы принципиально может дать начало любой клетке организма. Тело наземных растений — производное относительно немногих инициальных клеток.
Первичные меристемы обладают меристематической активностью, т. е. способны к делению изначально. В ряде случаев способность к активному делению может вновь возникнуть и у клеток, уже почти утративших это свойство. Такие «вновь» возникшие меристемы называют вторичными.
В теле растения меристемы занимают различное положение, что позволяет их классифицировать. По положению в растении выделяют верхушечные, или апикальные (от лат. «апекс» — верхушка), боковые, или латеральные (от лат. «латус» — бок), и интеркалярные меристемы.
Апикальные меристемы располагаются на верхушках осевых органов растения и обеспечивают рост тела в длину, а латеральные — преимущественно рост в толщину. Каждый побег и корень, а также зародышевый корешок, почечка зародыша имеют апикальную меристему. Апикальные меристемы первичны и образуют конусы нарастания корня и побега (рис. 1).
Латеральные меристемы располагаются параллельно боковым поверхностям осевых органов, образуя своего рода цилиндры, на поперечных срезах имеющие вид колец. Часть из них относится к первичным. Первичными меристемами являются прокамбий и перицикл, вторичными — камбий и феллоген.
Интеркалярные, или вставочные, меристемы чаще первичны и сохраняются в виде отдельных участков в зонах активного роста (например, у оснований междоузлии, в основаниях черешков листьев).
Существуют также раневые меристемы. Они образуются в местах повреждения тканей и органов и дают начало каллюсу — особой ткани, состоящей из однородных паренхимных клеток, прикрывающие место поражения Каллюсо-образовательная способность растений используется в практике садоводства при размножении их черенками и прививками. Чем интенсивнее каллюсообразование, тем больше гарантия срастания подвоя с привоем и укоренения черенков. Образование каллюса— необходимое условие культуры тканей растения на искусственных средах.
Клетки апикальных меристем более или менее изодиаметричны по размерам и многогранны по форме. Межклетников между ними нет, оболочки тонкие, содержат мало целлюлозы. Полость клетки заполнена густой цитоплазмой с относительно крупным ядром, занимающим центральное положение. Вакуоли многочисленные, мелкие, но под световым микроскопом обычно не заметны. Эргастические вещества, как правило, отсутствуют. Пластид и митохондрий мало и они мелки.
Клетки боковых меристем различны по величине и форме. Они примерно соответствуют клеткам тех постоянных тканей, которые из них в дальнейшем возникают. Так, в камбии встречаются как паренхимные, так и прозенхимные инициали. Из паренхимных инициалей образуется паренхима проводящих тканей, а из прозенхимных — проводящие элементы.

Рис. 1. Верхушечная меристема побега элодеи. А — продольный срез; 5 — конус нарастания (внешний вид и продольный срез); В — клетки первичной меристемы;
Г — паренхимная клетка сформировавшегося листа:
1 ~ конус нарастания, 2 — зачаток листа, 3 — бугорок пазушной почки
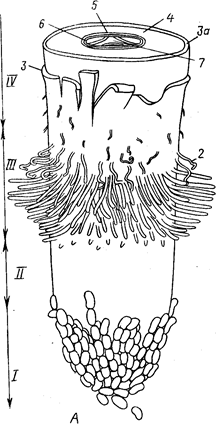
Зоны корня. Особенности морфологического строения каждой зоны в связи с выполняемыми функциями. Рост корня в длину и втолщину.
Наряду с побегом корень - основной орган высшего растения, в типичном случае выполняющий функцию минерального и водного питания. Другая важная функция корня, тесно связана с основной, - закрепление, растения в почве.
Различные части корня выполняют неодинаковые функции и характеризуются определенными морфологическими особенностями. Эти части получили название зон (рис. 2). Кончик корня снаружи всегда прикрыт корневым чехликом, защищающим апикальную меристему. Клетки корневого чехлика продуцируют слизь, покрывающую поверхность корня. Благодаря слизи снижается трение о почву, и ее частицы легко прилипают к корневым окончаниям и корневым волоскам.
Корневой чехлик состоит из живых паренхимных клеток, возникающих у большинства однодольных из особой меристемы, получившей название калиптрогена, а у двудольных и голосеменных — из верхушечной меристемы кончика корня. Водные растения корневого чехлика обычно не имеют.
Под чехликом располагается зона деления, представленная меристематической верхушкой корня, его апексом. В результате активности апикальной меристемы формируются все прочие зоны и ткани корня.
Делящиеся клетки сосредоточены в зоне деления, имеющей размеры около 1 мм. Эта часть корня заметно отличается от прочих зон своей желтоватой окраской. Вслед за зоной деления располагается зона растяжения (роста). Она также невелика по протяженности (несколько миллиметров), выделяется светлой окраской и как бы прозрачна. Клетки зоны роста практически не делятся, но способны растягиваться в продольном направлении, проталкивая корневое окончание в глубь почвы. Они характеризуются высоким тургором, что способствует активному раздвиганию частиц почвы. В пределах зоны роста происходит дифференциация первичных проводящих тканей.
Окончание зоны роста заметно по появлению на эпиблеме многочисленных корневых волосков. Корневые волоски располагаются в зоне—всасывания, функция которой понятна из ее названия. На корне она занимает участок от нескольких миллиметров до нескольких сантиметров. В отличие от зоны роста участки этой зоны уже не смещаются относительно частиц почвы. Основную массу воды и растворов солей молодые корни усваивают в зоне всасывания с помощью корневых волосков.
Корневые волоски появляются в виде небольших сосочков — выростов клеток эпиблемы. Рост волоска осуществляется у его верхушки. Оболочка корневого волоска растягивается быстро. По прошествии определенного времени корневой волосок отмирает. Продолжительность его жизни не превышает 10—20 дней.
Выше зоны всасывания, там, где исчезают корневые волоски, начинается зона проведения. Строение этой зоны на разных ее участках неодинаково. По этой части корня вода и растворы солей, поглощенные корневыми волосками, транспортируются в вышележащие отделы растения. Участки зоны проведения занимают фиксированное положение относительно участков почвы, не смещаясь относительно их. Несмотря на фиксированное положение в пространстве конкретных участков зон поглощения и проведения, эти зоны сдвигаются вследствие верхушечного роста. В результате этого всасывающий аппарат постоянно перемещаетсяв почве.
В пределах одной корневой системы существуют корни, выполняющие разные функции. У большинства растений различаются ростовые и сосущие окончания. Ростовые окончания долговечнее, относительно мощные, быстро удлиняются и продвигаются в глубь почвы. Сосущие окончания недолговечны, возникают в большом числе на вростовых корнях
и удлиняются медленно. У деревьев и кустарников выделяют видоизмененные скелетные и полускелетные корни, на которых возникают недолговечные корневые мочки, несущие множество сосущих окончаний.
 |
 |
Рис.2 Общий вид {А) и продольный срез (5) корневого окончания (схема). 1— корневой чехлик; 11 - зона роста и растяжения; ///— зона корневых волосков, или зона всасывания;IV — начало зоны проведения (в этой зоне также закладываются боковые корни):
1- заложившийся боковой корень, 2 — корневые волоски на эпиблеме, 3 — эпиблема, За — экзодерма, 4 — первичная кора,
5 — эндодерма, 6 — перицикл, 7 — осевой цилиндр
Факторы среды и процессы роста и развития. Фитогормоны. Фотопериодизм. Факторы среды и процессы роста и развития.
Внешние факторы, или факторы среды, также оказывают заметный эффект на рост и развитие. Рассмотрим здесь главнейшие факторы — свет, тепло и влагу. Свет оказывает глубокое влияние на внешнее строение растений, причем его действие многообразно. Свет влияет на дыхание и прорастание семян, образование корневищ и клубней, формирование цветков, на листопад, переход почек в состояние покоя. Растения, выращенные при отсутствии света (этиолированные растения), обгоняют в росте растения, выросшие на свету. Интенсивное освещение нередко усиливает процессы дифференцировки.
Для каждого растения существует свой температурный оптимум роста и развития. Температурные минимумы для роста и развития в среднем лежат в интервале 5 —15°С, оптимумы — при 25 — 35°С, максимумы — в пределах 45 — 55°С. Низкие и высокие температуры могут нарушать покой семян, почек и сделать возможным их прорастание и распускание. Образование цветков — это переход из вегетативного состояния в генеративное. Индуцирование, т. е. ускорение этого процесса холодом, называют яровизацией. Без процесса яровизации многие растения (свекла, репа, сельдерей, озимые злаки) не способны к цветению.
Огромное значение для роста, прежде всего в фазе растяжения, имеет обеспеченность водой. Недостаток воды влечет за собой мелкоклеточность, а следовательно, отставание в росте.
Фитогормоны — это химические факторы, вырабатываемые в крайне малых количествах, но способные давать значительный физиологический эффект. Фитогормоны, выработанные в одной части растения, транспортируются в другую часть, вызывая там соответствующие изменения в зависимости от генной модели воспринимающей клетки.
Известны три класса фитогормонов, действующих по преимуществу как стимуляторы: ауксины, гиббереллины и цитокинины. Два класса гормонов — абсцизовая кислота и этилен — оказывают в основном тормозящее действие. Механизм действия различных гормонов неодинаков.
Многие процессы обмена веществ, роста, развития и движения подвержены ритмическим колебаниям. Иногда эти колебания следуют смене дня и ночи (циркадные ритмы), иногда связаны с длиной дня (фотопериодизм). Пример ритмических движений — ночное закрывание цветков, опускание и продольное складывание листьев, раскрытых и приподнятых в дневное время. Эти движения связаны с неравномерным тургором. Во многих случаях эти процессы контролируются внутренней хронометрической системой — физиологическими часами, по-видимому, существующими у всех эукариотических организмов. У растений важнейшая функция физиологических часов — регистрация длины дня и вместе с тем времени года, что определяет переход к цветению или подготовку к зимнему покою (фотопериодизм).
Виды, растущие на севере (севернее 60°с.ш.), должны быть преимущественно длиннодневными, поскольку их короткий вегетационный период совпадает с продолжительной длиной дня. В средних широтах (35—40° с. ш.) встречаются растения как длиннодневные, так и короткодневные. Здесь весенние или осеннецветущие виды относятся к короткодневным, а цветущие в разгар лета — к длиннодневным.
Фотопериод имеет большое значение для характера распространения растений. В процессе естественного отбора у видов генетически закрепилась информация о длине дня своих местообитаний и об оптимальных сроках начала цветения, Даже у растений, размножающихся вегетативно, длина дня определяет соотношение между сезонными изменениями и накоплением запасных веществ. Виды, индифферентные к длине дня, являются потенциальными осмополитами. Сюда же относятся и виды, цветущие с ранней весны до поздней осени. Другие виды не могут выходить за пределы географической широты, определяющей их способность к цветению при соответствующей длине дня. Фотопериодизм важен и в практическом отношении, поскольку он определяет возможности продвижения южных растений на север, а северных — на юг.
СОЦВЕТИЯ. Определение соцветия. Классификация. Особенности строения цимоидных соцветий. Примеры. Биологическое значение соцветий.
Цветки могут располагаться поодиночке или группами. В тех случаях, когда они располагаются группами, образуются соцветия. Соцветие может быть определено как часть победа, побег или система видоизмененных побегов, несущих цветки.
Соцветия обычно более или менее ограничены от вегетативной части растения.
Биологический смысл возникновения соцветий — в возрастающей вероятности опыления цветков как анемофильных, так и энтомофильных растений. Несомненно, что насекомое за единицу времени посетит гораздо больше цветков, если они собраны в соцветия. Кроме того, цветки, собранные в соцветия, более заметны среди зелени листьев, нежели одиночные цветки. Многие поникающие соцветия легко раскачиваются под влиянием движения воздуха, способствуя тем самым рассеиванию пыльцы.
Соцветия свойственны громадному большинству цветковых растений. Обычно соцветия группируются близ верхней части растения на концах ветвей, но иногда, особенно у тропических деревьев, возникают на стволах и толстых ветвях. Такое явление известно под названием каулифлории (от лат. «каулис» — стебель, «флос» — цветок). В качестве примера можно привести шоколадное дерево (Т heobr ота сасао). Считается, что в условиях тропическою леса каулифлория делает цветки более доступными для насекомых-опылителей.
Закладываются соцветия внутри цветочных или смешанных почек. У многих растений (бузина, сирень, гиацинт и др.) соцветие возникает как единое целое в результате деятельности одной меристемы.
Любое соцветие имеет главную ось, или ось соцветия, и боковые оси, которые могут быть разветвлены в различной степени или неразветвлены. Конечные их ответвления - цветоножки — несут цветки. Оси соцветия делятся на узлы и междоузлия. На узлах осей соцветия располагаются листья и прицветники (рис. 3).
Соцветие несет видоизмененные или неизмененные листья. Сильно видоизмененные листья называют прицветниками или брактеями. Соцветие, несущее неизмененные ассимилирующие листья, является фрондозным, т. е. олисгвенным. У брактеозного соцветия в узлах распо- лагаются прицветники. Иногда вследствие полной редукции брактей соцветие становится эбрактеозным. У него прицветники отсутствуют. Соцветия могут быть резко отделены от вегетативной части либо (особенно в случае фрондозных соцветий) эта граница выражена неясно.
Точная характеристика соцветий необходима во многих случаях, в том числе и при анализе морфологических особенностей лекарственных растений. Поэтому классификации соцветий уделяется значительное внимание.
Соцветия, у которых боковые оси ветвятся, называются сложными. У простых соцветий боковые оси не разветвлены и являются цветоножками. У сложного соцветия боковые оси несут частные, или парциальные, соцветия.
Существует мнение, согласно которому сложные соцветия — более древний тип. Во многих случаях простые соцветия возникали в процессе «обеднения» сложных, что связано с редукцией их боковых осей. Полагают, что одиночные цветки в пазухах листьев или на верхушках побегов также возникли в результате крайней редукции соцветий разного типа. Главная ось может заканчиваться верхушечным цветком; в этом случае соцветие ограничено в росте и получило название закрытого.
У открытых соцветий главная ось обладает неограниченным ростом и цветки располагаются сбоку от морфологической верхушки. У сложных соцветий верхушечными цветками могут заканчиваться главная и боковые оси или все они имеют неограниченный рост.
У обоеполых растений соцветия несут обоеполые цветки, но у однодомных и двудомных соцветия могут быть также тычиночными, пестичными и полигамными. В последнем случае встречаются одновременно тычиночные, пестичные и обоеполые цветки.
Классификация соцветий может осуществляться на основе особенностей ветвления конечных парциальных соцветий. В соответствии с этим соцветия подразделяют на два главных типа: ботриоидные и цимоидные. У ботриоидных соцветий (от греч. «ботрион» — кисть) характер ветвления моноподиальный. Цимоидные соцветия (от греч. «кюма» — волна по особом порядку зацветания) обязательно характеризуются симподиальным ветвлением парциальных соцветий.

Рис. 3. Строение соцветия:
1 — главная ось, 2 — боковая ось (паракладии), 3 — узлы, 4 — междоузлия, 5 — прицветники, 6 — цветоножки, 7 — цветки
 . Рис. 4. Типы соцветии А —
простые ботриоидные:
. Рис. 4. Типы соцветии А —
простые ботриоидные:
1-кисть, 2 — колос, 3 — початок, 4 — простой зонтик, 5 - головка, 6 — корзинка, 7 — щиток (4. 5, 6 — с укороченной главной осью, прочие — с удлиненной);
Б - сложные ботриоидные. Метелка и ее производные :1 - метелка, 2 — сложный щиток, 3 — антела;
В - сложные ботриоидные. Сложная кисть и ее производные:
1 - тройная кисть, 2 — двойная кисть, 3 — двойной колос, 4 — двойной зонтик
ЦИМОИДНЫЕ СОЦВЕТИЯ
Это обширная группа соцветий, встречающаяся так же часто, как и ботриоидные. Среди цимоидных соцветий выделяют два основных типа: цимоиды и тирсы. Цимоиды, как правило, представляют собой упрощенные тирсы. У всех цимоидных соцветий парциальные соцветия образуются за счет симподиального ветвления.
Различают три типа цимоидов: монохазии, дихазии и плейохази. У монохазиев под цветком, завершающим главную ось, развивается только одно парциальное соцветие или в простейших случаях единственный цветок. В соответствии с особенностями ветвления парциальных соцветий типа монохазиев принято выделять извилину, завиток и клубок. Монохазий довольно обычен у части лютиковых, в частности у лютика едкого ( R апипсиlus ас ris ). Парциальные соцветия в виде завитка встречаются у большинства представителей семейства бурачниковых. От главной оси дихазия, под завершающим ее цветком, отходят два парциальных соцветия, а в простейших случаях — два цветка. Возможны простые, двойные, тройные дихазии и т. д. Диха-зий встречается у ряда гвоздичных, например видов рода звездчатка ( Stellaria)
Плейохазии характеризуются строением, при котором под цветком, завершающим главную ось, развиваются три или более парциальных соцветия (или цветка). Принципиально возможны двойные, тройные и т. д. плейохазии (рис. 5). Для рода молочай (Еир horbi а) из семейства молочайных характерен особый тип цимоидного соцветия, получивший название циатий. Циатий состоит из верхушечного пестичного цветка и пяти тычинок, возникших вследствие крайней редукции пяти тычиночных парциальных соцветий. Циатий окружен обверткой, состоящей из листьев редуцированных парциальных соцветий. Тирсы устроены сложнес, чем цимоиды. Это разветвленные соцветия, причем степень разветвления уменьшается от основания к верхушке. Главная ось тирса нарастает моноподиально, но парциальными соцветиями того или иного порядка являются цимоиды.
Возможны различные классификации тирсов. Тирсы, главная ось которых заканчивается цветком, называются закрытыми, в противном случае они считаются открытыми. В зависимости от степени разветвления боковых осей выделяют плейотирсы, у которых цимоиды размещаются на осях третьего и более высоких порядков; дитирсы, у которых цимоиды расположены на осях второго порядка, и монотирсы, у которых цимоиды находятся непосредственно на главной оси соцветия (см. рис. 4).
Внешнее сходство тирсов с кистью, колосом, сережкой, зонтиком или головкой позволяет говорить о кистевидном, колосовидном, сережковидном, зонтиковидном, головковидном тирсах и т. п. Тирсы легко преобразуются в другие типы соцветий в ходе редукции (уменьшения) числа боковых осей, укорочения междоузлии и т. д. В результате редукции возникают цимоиды — соцветия, по внешнему виду подобные ботриоидным, и даже одиночные цветки.
Тирсы встречаются у растений очень часто. Например, тирс — соцветие конского каштана (Ае s си lus hippocastanum ), другой пример тирса — соцветие коровяка (Verbascum) из семейства норичниковых. Тирсы различных типов представляют собой соцветия всех губоцветных. Соцветие березы — сережковидный тирс.
ИСПОЛЬЗОВАНИЕ ЦВЕТКОВ И СОЦВЕТИЙ
Цветки имеют разнообразное практическое применение. Очень часто красотой определяется их декоративное использование и эстетическое воздействие на человека. Растения с цветками привлекательной окраски, формы или запаха широко культивируются во множестве искусственно выведенных сортов. Известны, например, около 25 тыс. сортов роз, около 4 тыс. сортов тюльпанов, 12 тыс. сортов нарциссов и т. д. Учреждены специальные общества любителей и коллекционеров тюльпанов, лилий, орхидей, роз. В России и за рубежом существуют фирмы, специализирующиеся на разведении и продаже красивоцветущих растений. Особенно высоко поставлена культура декоративных луковичных однодольных (тюльпаны, гиацинты) в Нидерландах. В Сингапуре и ряде стран Юго-Восточной Азии в промышленных масштабах выращивают орхидеи. Срезанные цветки широко используют в виде букетов, гирлянд, венков и т. д. В Японии искусство составления букетов получило название икебана.
Многие цветки содержат ароматические эфирные масла. Одно из известнейших в Европе розовое масло, получаемое из лепестков махровых форм розы дамасской ( Rosa damascena ), розы столистной (Rosa centifolia) и некоторых других видов, применяется в парфюмерии и отчасти в медицине. В медицине употребляют много цветков различных растений. Особенно популярны цветки ромашки аптечной (Ма tri са ri а r еси tit а), цветки липы (Т ili а со rd а t а), цветки календулы, или ноготков (Са1еп d и1а о ffici па lis ), бутоны софоры японской ( St ур h по1о biu т japonicum) — источника промышленного получения витамина Р (рутина) и др.
Ряд цветков применяют для ароматизации вин и Табаков. Из них особенно известен донник лекарственный (Ме lil о tus о fficinalis ), чьи цветки, содержащие кумарин, добавляют во многие сорта табака. Пищевые красители получают из цветков уже упоминавшихся ноготков лекарственных, рылец культивируемого шафрана посевного (С r ос us s а tivus ) и сафлора красильного (Са rthamus tinctorius ). Зеленые бутоны каперсов колючих (Сарра ris spinosa ) маринуют в качестве острой приправы. Ценную пряность представляют бутоны сизигиума ароматного ( Syzygium а r ота ti сит). Они известны под названием гвоздики, что связано с формой высушенных бутонов. В тропиках многие крупные цветки используют как овощ. Довольно широкое применение в этом качестве находят бутоны цветков некоторых видов бананов (М usa ). Нектар цветков служит источником получения разных сортов меда. Наиболее ценные медоносы — разные виды липы, фацелия пижмолистная (Р h асе li а t апасе tifolia ) и гречиха съедобная ( F а g ору r ит е s си1еп t ит). В последние десятилетия в медицине стали использовать препараты из пыльцы растений.
Рис. 4. Агрегатные соцветия:
1 — метелка зонтиков, 2 — метелка корзинок, 3 — щиток корзинок, 4 — кисть корзинок, 5— колос корзинок

Рис. 5. Цимоидные соцветия. А — цимоиды:
1
—3 —
монохазии: /—«элементарный» монохазий, 2 —
извилина, 3—завиток, 4—
двойной завиток, 5—6—
дихазии: 5—дихазии, 6—тройной дихазии, 7—8—
плейохазии: 7—плейохазии, 8—
двойной плейохазии;  Б —
пример тирса
Б —
пример тирса
Анатомическое строение корнеплодов (видоизменений корня), типа редьки, моркови и свеклы. Особенности строения и расположения тканей в корнеплодах.
Наряду с функциями поглощения и проведения корни выполняют функцию запаса питательных веществ, которые иногда накапливаются в очень больших количествах. Таковы корни некоторых двулетних двудольных растений (морковь, петрушка, репа, редька, свекла и др.), образующие корнеплоды.
В состав корнеплода кроме корня входит подсемядольное колено (гипокотиль), а также морфологически нижняя часть побега. У разных «корнеплодов» эти органы развиты и утолщены в разной степени. Так, у моркови, петрушки, некоторых сортов редьки и сахарной свеклы большая часть «корнеплода» образованна главным корнем; у репы, столовых сортов свеклы и других – основанием стебля и гипокотилем, а собственно корень представляет собой лишь самую нижнюю часть «корнеплода», несущую боковые корни.
Запасные вещества, отложенные в «корнеплоде» в первый год жизни растения, на следующий год расходуются на развитие цветоносных побегов, плодов и семян.
Питательные вещества накапливаются в живых паренхимных клетках с тонкими неодревесневшими стенками. Клетки богаты клеточным соком, поэтому корнеплоды всегда сочные.
Вещества могут откладываться: 1) преимущественно в древесной (кселимной) паренхиме (редька, репа и другие крестоцветные); 2) в древесной и особенно в лубяной (флоэмной) паренхиме (морковь, петрушка и другие зонтичные); 3) в паренхиме, образованной деятельностью нескольких добавочных камбиев (свекла).
Корень моркови (Daucus sativus (Hoffm.) Roehl.)
На поперечном срезе, еще не обработанном реактивами, хорошо различимы две зоны: внутренняя, довольно узкая, светло-желтая (вторичная ксилема) и наружная – более широкая , оранжевая (вторичная флоэма). Во флоэме сосредоточенна основная масса крахмала, растворимых сахаров и других питательных веществ. Оранжевый цвет обусловлен присутствием в ее клетках хромопластов с кристаллами каротина. Флоэма и ксилема разделены светлой камбиальной зоной, которая часто разрывается, и проводящие ткани отделяются одна от другой.
На срезе, обработанном флороглюцином и соляной кислотой, уже при малом увеличении микроскопа можно обнаружить два очень коротких, узких, обычно сходящихся в центре корня тяжа трахеальных элементов первичной кселимы, окрасившихся в красный цвет. От наружных концов каждого из тяжей этой диархной ксилемы к периферии среза отходят крупноклеточные первичные лучи. Между ними, по обе стороны от первичной ксилемы, находится вторичная ксилема, пересеченная многочисленными вторичными лучами (рис.6). Основная масса ксилемы состоит из тонкостенных паренхимных клеток. Сосудов в корне моркови немного. Они резко выделяются на общем фоне бесцветной паренхимы, так как имеют утолщенные одревесневшие стенки. Сосуды расположены поодиночке или собраны по три – семь в группы. Во внутренних, наиболее старых участках вторичной кселемы они составляют прерывистые радиальные полосы. Паренхимные клетки периодически делятся в разных направлениях, поэтому в толстых корнях все элементы сильно смещены.
Камбиальная зона обычно широкая, клетки ее мелкие, сжатые в радиальном направлении.
Флоэма состоит главным образом из паренхимных элементов. Среди них находятся небольшие группы ситовидных трубок с сопровождающими клетками и схизогенные эфирномасляные каналы. На срезе хорошо заметны лучи из очень крупных клеток. Периодические деления паренхимных клеток обусловливают значительное разрастание флоэмы в толщину. Наружная часть флоэмы граничит с зоной из нескольких (6-7) рядов крупных паренхимных клеток, возникших, вероятно, вследствие делений клеток феллодермы. В этой зоне видны эфирномасляные каналы. Корень покрыт тонким слоем пробки.
Корень редьки (Raphanus sativus L.)
В отличие от моркови утолщение корня редьки происходит главным образом за счет сильного разрастания вторичной кселимы. Вторичная флоэма по сравнению с мощной ксилемой развита весьма слабо.
В центре поперечного среза (рис.7) находятся мелкие сосуды диархной первичной ксилемы. От каждого ее конца, пересекая в радиальном направлении центральный целиндр, отходят по одному широкому первичному паренхимному лучу.
Запасающая ткань корня представлена вторичной ксилемой, основная масса которой сложена тонкостенными клетками тяжевой и лучевой паренхимы; клетки ее содержат крахмал и богатыклеточным соком. Немногочисленные широкопросветные сосуды собраны по два – шесть в короткие радиальные цепочки, расширяющиеся к периферии. Вблизи камбия сосуды обычно окружены небольшим числом механических элементов со слабоутолщенными, иногда одревесневшими стенками.
Камбиальная зона состоит из мелких таблитчатых клеток с густым цитоплазматическим содержимым. Снаружи она окружена узким кольцом вторичной флоэмы, в которой, как и в ксилеме, заметны широкие вторичные лучи. Вытянутые в радиальном направлении группы ситовидных трубок с сопровождающими клетками окружены паренхимой. К концу вегетации во вторичной флоэме выявляются тяжи коротких волокон. Их слабоутолщенные одревесневшие стенки имеют немногочисленные простые поры. Вокруг флоэмы расположен тонкий слой паренхимных клеток - производных перицикла и феллодермы. Корень покрыт бурой пробкой.
Корень свеклы ( Beta vulgaris L.)
На поперечном срезе корня обращают на себя внимание чередующиеся концентрические кольца более или менее интенсивно окрашенных тканей (рис.8).
Начальные стадии утолщения корня свеклы сходны с утолщением корней моркови и редьки, но в дальнейшем его разрастание в толщину происходит своеобразно.
Корень проростка в фазе семядольных листьев имеет первичное строение. Радиальный проводящий пучок с диархной первичной ксилемой окружен однослойным перециклом. С появлением у растения первого настоящего листа начинает функционировать камбий, заложившийся между первичной ксилемой и флоэмой. В результате его деятельности на 12-15-й день жизни сеянца в центре корня по обеим сторонам от первичной ксилемы образуется по одному пучку, состоящему из вторичных ксилемы и флоэмы. Сильно облитерированная к этому времени первичная флоэма оттесняется к периферии. Вторичная ксилема состоит из сосудов с одревесневшими стенками и небольшого числа паренхимных клеток. Между пучками вторичных тканей проходят широкие лучи. Деятельность этого первичного камбия вскоре заканчивается и дальнейшее утолщение корня продолжается за счет добавочных камбиев, возникающих последовательно один за другим при непосредственном участии перицикла, делясь преимущественно тангентальными перегородками, образуют многослойную меристематическую ткань, которая на поперечном срезе расположена кольцом. В периферической части этого кольца закладывается феллоген, клетки средней части дифференцируются в паренхимные элементы, а из внутреннего ряда клеток формируется первый добавочный камбий, образующий новую зону меристиматических клеток. Наружный ряд клеток этой зоны в дальнейшем будет функционировать в качестве второгодобавочного камбия. Внутренние клетки меристематической зоны, делясь, откладывают наружу тонкостенные клетки паренхимы и небольшие группы клеток вторичной флоэмы, внутрь – крупные клетки перенхимы и непосредственно под участками флоэмы – элементы кселемы.
Таким образом, в результате деятельности первого добавочного камбия возникает широкое кольцо паренхимной ткани с погруженнымит в нее мелкими коллатеральными пучками. Флоэма состоит из нескольких ситовидных трубок с сопровождающими клетками и клеток паренхимы, ксилема – из небольшого числа пористых сосудов, окруженных механическими элементами, и ксилемной паренхимы (рис 8, Б).
Вскоре начинают делится клетки второго добавочного камбия, образуя следующую меристиматическую зону. Наружные клетки ее дифференцируются в третий добавочный камбий, а внутренние формируют второе кольцо проводящих пучков и межпучковую паренхиму. Третий добавочный камбий в свою очередь участвует в образовании четвертого добавочного амбия и третьего кольца проводящих пучков и т.д.
Деятельностью добавочных камбиев объясняется наличие на поперечном срезе корня нескольких (8 и более) концентрических колец, состоящих из мелких проводящих пучков и обильной паренхимы, в тонкостенных клетках которой накапливаются сахароза и другие питательные вещества. У столовых сортов свеклы в паренхимных клетках много антоциана. Чем более удалены добавочные камбии от центрва корня, тем слабее их деятельность. Поэтому ширина концентрических колец, число и размеры проводящих пучков уменьшаются к перифериисреза. Самые молодые наружные кольца могут быть представлены лишь узкими слоями слабодифференцированных или даже меристиматических клеток.
Интенсивность вторичного утолщения корня свеклы тесно связанна с развитием листового аппарата. Установлено, что число концентрических слоев зависит от числа листьев в прикорневой розетке.
Деятельность добавочных камбиев заканчивается рано, и дальнейшее утолщение корня происходит за счет деления и разрастания паренхимных клеток.
Снаружи корня находится тонкий слой мелкоклеточной паренхимной ткани, окруженной темно-бурой пробкой.
Похожие рефераты:
Общее содержание воды в листьях калины в условиях биостанции
Влияние 6-БАП на растения кукурузы при разном уровне засоления
Анатомическое строение растений
Физиология и питание растительных организмов
Программа обучения + тесты по биологии для школы.zip
Методика изучения ампельных растений на уроках биологии
Методические особенности изучения темы "Корень" в школьном курсе биологии
Формирование умений и навыков у учащихся второго класса при изучении комнатных растений
Разнообразие декоративных растений
Взаимодействие микроорганизмов с высшими растениями
Строение растительной клетки. Ткани растений
Декоративное озеленение интерьера
Экологическая оценка состояния популяции редкого вида Касатика (Ириса) карликового