| Скачать .docx | Скачать .pdf |
Реферат: Происхождение эукариотических клеток
КУРСОВАЯ РАБОТА
по биологии
тема:
“Происхождение эукариотических
клеток”
Выполнила: Сурова Вера
10-В класс,школа N55
Проверила: Струкова Н.А.
г.Рязань 1999 год
ПЛАН
1 . Симбиотическая теория :
1.1 Фундаментальное разграничение между прокариотами и эукариотами ;
1.2 Прокариотические организмы ;
1.3 Эукариотические клетки ;
1.4 Митохондрии и пластиды ;
1.5 Способность эукариот к фотосинтезу ;
2. Предшественники жизни ;
3. Ферментирующие организмы :
3.1 Источники энергии ранних клеток ;
3.2 Метаногенез как источник энергии ;
3.3 Цианобактерии и хлорооксибактерии ;
4. Фотосинтез и воздух ;
5. Ассоциации и эукариоты :
5.1 Теории последовательных симбиозов ;
5.2 Симбиотическое приобретение подвижности ;
5.3 Эукариоты – фотосинтетики ;
6. Взгляд на эволюцию клетки :
6.1 Гипотеза прямой филиации ;
6.2 Гипотеза : от цианобактерий к водорослям ;
6.3 Совместимость 2 –х гипотез : прямой филиации и
эндосимбиоза ;
7. Выводы ;
8. Литература .
ТЕОРИЯ ЭНДОСИМБИОЗА
Все живые организмы на Земле обычно делят на прокари-от и эукариот . Главной особенностью прокариот в отличие от эукариот является отсутствие у них полноценного клеточного ядра , покрытого мембраной . Клетки прокариот имеют очень не-большие размеры порядка 1 мкм . Объем эукариотических клеток, содержащих полноценное ядро, в 800 – 1000 раз больше объема клеток прокариот . В связи с этим клеткам – эукариотам необходим для выполнения жизненных функций гораздо больший мембранный аппарат . Действительно , в эукариотической клетке выявлены субклеточные структуры , окруженные незави-симыми от плазмалеммы и друг от друга мембранами . Эти структуры называются органеллами . Каждой органелле : митохондрии , хлоропласту , ядру , лизосомам , аппарату Гольджи и т. д . присущи свои специфические функции в жизни клетки .
С помощью молекулярно – биологических подходов получены убедительные доказательства того ,что все живые существа должны быть разделены на 3 надцарства : бактерии , археи и эукариоты . Бактерии и археи возникли на Земле от общего предка - так называемого прогенота - около 4 млрд лет назад , а эукариоты по данным палеонтологов появились на Земле через 500 млн лет после прокариот . Наиболее популярной и достаточно обоснованной идеей происхождения эукариотических клеток , в настоящее время являктся идея многократного эндосимбиоза различных прокариот . По представлениям известных микробиологов А. Клюйвера и К. Ван Ниля , именно у древних прокариот произошел отбор клеток с наиболее экономичными и зачастую полифункциональными биохи-мическими реакциями , ставшими основой важнейших путей метаболизма .
Симбиотическая теория происхождения и эволюции клеток основана на двух концепциях , новых для биологии .Согласно первой из этих концепций, самое фундаментальное разграни-чение в живой природе - это разграничение между прокариотами и эукариотами ,т.е. между бактериями и организмами, состоящими из клеток с истинными ядрами - протистами , животными , грибами и растениями . Вторая концепция состоит в том , что источником некоторых частей эукариотических клеток была эволюция симбиозов , т.е. формирование постоянных ассоциаций между организмами разных видов . Предполагается , что три класса органелл - митохондрии , реснички и фотосинтезирующие пластиды – произошли от свободно живущих бактерий , которые в результате симбиоза были в определенной последовательности включены в состав клеток прокариот - хозяев . Эта теория в большой мере опирается на неодарвинистские представления , развитые генетиками, экологами , цитологами и другими учеными , которые связали Менделевскую генетику с дар-виновской идеей естественного отбора .Она опирается также на совершенно новые или недавно возрожденные научные направ-ления : на молекулярную биологию ,особенно на данные о структуре белков и последовательности аминокислот , на микро палеонтологию , изучающую наиболее ранние следы жизни на Земле , и даже на физику и химию атмосферы , поскольку эти науки имеют отношение к газам биологического происхсждения . Все организмы, состоящие из клеток, могут быть сгруппиро-ваны в пять царств : царство прокариот (Monera, куда относятся бактерии ) и четыре царства эукариот (Protoctista, Animalia, Fungi , Plantae). Протоктисты- это эукариотические организмы, не относящиеся к животным, грибам или растениям. В царство протоктистов входят водоросли , протозои, слизевики и другие эукариотические организмы неясной принадлежности . Протисты определены более ограничительно- как однокле-точные эукариоты . Таким образом , царство Protoctista включает не только протистов – одноклеточных эукариот , но и их ближай-ших многоклеточных потомков, таких как красные и бурые водоросли ,а также многие микроорганизмы ,сходные с грибами ,например хитридиевые . Согласно традиционному представ-лению о прямой филиации , такие клеточные органеллы , как митохондрии и пластиды , возникли путем компартментализации самой клетки .
НЕСКОЛЬКО ПРОКАРИОТ ОБРАЗУЮТ
ОДНУ ЭУКАРИОТУ
Клетки - это ограниченные мембранами системы , наименьшие из биологических единиц , способных к саморепродукции . Даже мельчайшие клетки содержат гены в форме молекул ДНК (одной или нескольких ) и белоксинтезирующий аппарат , состоящий из нескольких типов РНК и множества белков. Все клетки содержат рибосомы – тельца диаметром около 0,02 мкм , состоящие из РНК по меньшей мере трех типов , и примерно пятидесяти различных белков. Прокариотичесие организмы – наименьшие биологические единицы , которые удовлетворяют этому определению клетки . У них нет ядер . С другой стороны , одиночные эукариотические ( имеющие ядро ) клетки в известном смысле вообще не являются единицами . Это комплексы различных белоксинтезирующих единиц ( ну-клеоцитоплазмы , митохондрий и пластид ) , тип и число которых варьирует в зависимости от видовой принадлежности . Таким образом , согласно этой теории , эукариотические клетки возникли в результате кооперации первоначально независимых элементов, объединившихся в определенном порядке . Тэйлор назвал такое представление о раздельном происхождении и развитии частей эукариотической клетки и их последующем объединении - теорией последовательных эндосимбиозов . Условия высокой температуры и кислотности привели к выработке в нуклеоцитоплазме особого класса белков - гистонов - для защиты ее ДНК . Митохондрии были способны расщеплять трехуглеродные соединения до углекислоты и воды . Симбиотическая ассоци-ация нуклеоцитоплазмы и митохондрий начала изменяться в сторону усиления их взаимной зависимости . Этот комплекс включил в себя двигательные органеллы - реснички и жгутики . За этой трансформацией последовало морфогенетическое преобразование , которое состояло в развитии выраженных клеточных асимметрий . С интеграцией взаимозависимых нуклеоцитоплазмы, митохондрий, ундулиподий - завершилось создание полигеномной структуры эукариотической клетки. Первая эукариотическая клетка была гетеротрофной. Она питалась готовыми органическими соединениями. У этих эукариот возникли процессы митоза, а затем и мейоза. И наконец , приобретение эукариотами способности к фотосинтезу , имело место в процессе становления основной эукариотической организации и после его завершения- последнее событие в зтой эпопее. Симбиозы становились более и более интегрированными . В настоящее время зависимость каждой органеллы от продуктов метаболизма других органелл стала настолько полной , что только современные методы анализа позволяют проследить метаболические пути исходных партнеров . Эукариоты разнообразны по своему строению , но метаболически они остаются единообразными . Метаболические ухищрения эукариот были разработаны различными бактериями еще до того времени , когда они объединились, превратившись в хозяина и его органеллы . Прокариоты достигли большого разнообразия в способах получения энергии и в тонкостях биосинтеза . Они оставили след своего существования задолго до того , как на основе эукариотической клетки сформировались такие крупные организмы , как животные , обладающие скелетом .
Схема про исхождения эукариотических клеток путем симбиоза
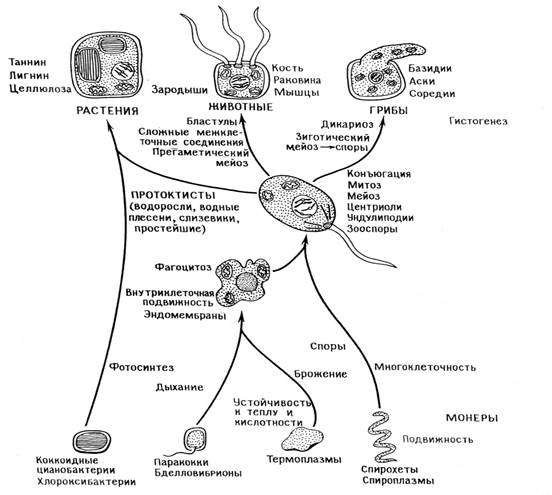
ПРЕДШЕСТВЕННИКИ ЖИЗНИ
Предки нынешних организмов возникли более трех милли-ардов лет назад из органических соединений , образовавшихся на поверхности Земли . Привлекательность этой концепции отчасти связана с ее доступностью для проверки .
С. Миллер и Х.. Юри показали , что при атмосферных усло-виях , имевших место на юной Земле , могут образовываться аминокислоты и другие органические молекулы . Различные типы органических молекул образовывались спонтанно в течение первого миллиарда лет существования Земли . Многие органические молекулы были обнаружены в межзвездном пространстве и в метеоритах . Значит , органические соединения возникают при взаимодействии легких элементов, в при-сутствии источников энергии . Универсальность химического механизма биорепродукции означает , что все живое на Земле происходит от общих предков : вся нынешняя жизнь произошла от клеток с информационными системами , осно-ванными на репликации ДНК , и на синтезе белков , направляемом информационной РНК .
ФЕРМЕНТИРУЮЩИЕ ОРГАНИЗМЫ
Возможно , что нуклеиновые кислоты с самого начала были отделены от окружающей среды липопротеидными мембранами . Информация стала кодироваться последовательностью нуклеотидов в молекулах ДНК . Древнейшие клетки использовали АТФ и аминокислоты , входящие в состав белков , как прямые источники энергии , и как структурные компоненты . Итак - древнейшая жизнь существовала в форме гетеротрофных бактерий , получавших пищу и энергию из органического материала абиотического происхождения . Для неодарвиновской органической эволюции необходимо три феномена : воспроизведение, мутации и селективное давление среды . Воспроизведение - это неприменное условие биологи-ческой эволюции . При достаточно точной репродукции - мутации и естественный отбор неизбежны . Объяснить проис-хождение такой репродукции - значит понять происхождение самой жизни . Мутирование - первичный источник изменчивости в эволюции , ведет к наследуемым изменениям в самовоспроизводяйщейся системе . Как только выработалось надежное воспроизведение , началась неодарвиновская эволю-ция . Организмы , у которых выработались пути синтеза нужных клетке компонентов , имели преимущество перед другими . Древнейшими автотрофами , то есть организмами способными удовлетворять свои потребности в энергии и органических веществах за счет неорганических источников , были анаэробные метанообразующие бактерии . Способность к фотосинтезу впервые выработалась у бактерий , чувствительных к кислороду . Поглощая солнечный свет с помощью бактериального хлорофилла , они превращали угле-кислоту атмосферы в органические вещества .Появление ана-эробного фотосинтеза необратимо изменило поверхность и атмосферу Земли .
Углекислота удалялась из атмосферы и превращалась в ор-ганическое вещество в соответствии с общим уравнением фотосинтеза : 2H2X+CO2=CH2O+2X+H2O . Фотосинтезирующие бактерии , которые использовали для восстановления СО2 сероводород , откладывали элементарную серу . Они дали начало бактериям , способным к фотосинтезу с выделением кислорода . Вероятно этот тип фотосинтеза зародился в результате накопления мутаций у фотосинтезирующих серных бактерий . Этот путь привел к возникновению сине-зеленых водорослей или цианобактерий . Недавно была открыта и изучена новая группа фотосинтезирующих прокариот , выделяющих О2 - хлороксибактерии . Можно предполагать , что именно циано- и хлорооксибактерии были предками пластид , находящихся в клетках водорослей и растений . По мере накопления О2 в результате микробного фотосинтеза земная атмосфера становилась окислительной , что вызвало кризис всей жизни – до этого времени мир был заселен анаэробными бактериями. Чтобы выжить микробы должны были избегать кислорода, укрываясь в болотах , или должны были выработать обмен веществ , способный предохранить их от возрастающего количества кислорода .
ФОТОСИНТЕЗ И ВОЗДУХ
Примерно 2 миллиарда лет назад один за другим происходили различные эволюционные процессы по созданию метаболи-ческого разнообразия . Способность к выработке специфических ферментов может передаваться в пределах одного вида с помощью вирусоподобных частиц, называемых плазмидами. Поразительны биосинтетические способности прокариот : из углекислоты , нескольких солей и восстановленных неорга-нических соединений , служащих источниками энергии , микро-организмы- хемоавтотрофы могут синтезировать в содержащей кислород неосвещенной воде все сложнейшие макромолекулы , необходимые для роста и размножения . Эти биохимические про-цессы приводили к системам круговорота элементов еще тогда , когда не было растений , грибов , животных .
Ассоциации и эукариоты .
По мере накопления атмосферного кислорода абиотическая продукция органических веществ сокращалась , так как они быстро разрушались под действием этого газа . Бактерии оказались в прямой зависимости друг от друга о том , что касается притока газов , вентиляции и удаления отходов мета-болизма в газообразном виде . Создалась их взаимная зависи-мость в отношении питания и защиты , возникло много разного рода связей между организмами , включая симбиоз , паразитизм и хищничество. Определенная серия симбиозов вела к форми-рованию новых типов клеток . Согласно теории последова-тельных симбиозов , все эукариоты сформировались в резуль-тате симбиоза между чрезвычайно далекими друг от друга видами прокариот : нуклеоцитоплазма образовалась из микро- организмов хозяев , митохондрии из бактерий , дышащих кислородом , пластиды - из хлороокси- или цианобактерий , а ундолиподии - из спирохет , прикреплявшихся к поверхности хозяев . Митотическое деление клетки выработалось только после того , как хозяева стали поглощать спирохет и передислоци-ровать их части . Итак , в начале истории эукариот находились лишенные клеточной стенки плейоморфные микроорганизмы , способные сбраживать глюкозу до двух- и трех углеродных ко-нечных продуктов по метаболическому пути Эмбдена-Мейер-гофа. Эти организмы , которым предстояло стать нуклео-цитоплазмой , приобретали эндосимбионтов . А какова природа эндосимбионтов , дышавших кислородом ? Эти промитохондрии были аэробными палочковидными организмами , близкими к современным бактериям , вроде Paracoccus denitrificans. Ассоциация могла начаться с хищничества , объектом которого служили клетки-хозяева . Размножение симбионтов внутри ос-мотически регулируемой клетки вело к отбору, направленному против сохранения ими клеточных стенок и биосинтетических путей, дублирующих пути хозяина. Естественный отбор привел к образованию крист - складчатых мембран современных митохондрий . Итогом этого приспособления к симбиозу было образование двугеномных клеток – аэробных , содержащих митохондрии и ядра , но неспособных к митозу , крупных амебоидных микроорганизмов . Следующим шагом к полному эукариотическому статусу было симбиотическое приобретение подвижности . Ундулиподии образовались только после того , как спирохеты объединились с амебоидами и навсегда видоизменились . Как и во всех устойчивых ассоциациях , репродукция симбионтов стала протекать согласованно с репродукцией хозяев и появились новые комплексы , содер-жащие три генома : ядерный, митохондриальный и ундолипо-диальный . Эти тригеномные комплексы дали множество различных видов протистов . Симбиотические бактерии став-шие ундолиподиями , обеспечили преадаптацию к митозу . Развитие механизмов митоза привело в результате адаптивной радиации к образованию огромного множества видов эукарио-тических микроорганизмов . Итак , гетеротрофные микробы – эукариоты породили животных и грибы . Другая серия симбиотических приобретений привела к эукариотам - фотосинтетикам . Естественный отбор оптимизировал процес-сы митоза, мейоза . Протопластиды начали ассоциироваться с гетеротрофными микрооорганизмами , которые заглатывали микробов – фотосинтетиков , но не переваривали их . Прокариоты хлороксибактерии - прародители пластид простей-ших , зеленых водорослей . Цианобактерии - стали предками красноватых пластид красных водорослей и криптомонат. Великие новшества в эволюции клеток возникли еще до того , как появились первые животные растения и грибы . Какие – то гетеротрофные эукариоты в результате поглощения фотосинтезирующих прокариот стали водорослями . Это завершающее событие произошло более 400 млн. лет назад , о чем свидетельствуют ордовикские ископаемые остатки крупных фотосинтетиков - сифоновые зеленые водоросли и красные водоросли .
Взгляд на эволюцию клетки.
Прямая филиация.
Традиционный взгляд на происхождение и эволюцию эукарио-тических клеток заключается в том , что все живые организмы произошли прямым путем от единственной предковой популя-ции в результате накопления мутаций под действием естест-венного отбора.
Точечные мутации, дупликации, делеции и другие наследствен-ные изменения , как известно игравшие роль в эволюции живот-ных и растений , ответственны и за дифференцировку эукарио-тических клеток из прокариотических- теория прямой филиации . т. е. представление о несимбиотическом происхождении клеток. Возможно , зеленые водоросли являются потомками родоначаль-ников всех растений. Многие особенности этих водорослей наводят на мыысль , что они и растения имеют общего предка. Филогения животных сложнее , но концепция прямой филиации вероятно приложима также и к их путям эволюции . Губки очевидно произошли от протистов , независимо от остальных групп животных . Итак , по крайней мере для животных и растений гипотеза , по которой сложные эукариотические орга-низмы развились из более просто устроенных эукариот путем накопления отдельных мутаций под действием естественного отбора , вполне правомерна . Эволюционные новшества опирают-ся на сохранение высокосовершенных благоприятных генов . Итак , новые организмы с повышенной способностью контроли-ровать свою среду обитания развивались больше благодаря сохранению дупликации и рекомбинации высокоадаптированных генов ,чем путем приобретения новых свойств в результате простых , случайных точечных мутаций . Гипотеза прямой филиации не позволяет объяснить происхождение эукариот от прокариот . Например хиатус между неспособными к митозу цианобактериями , и такими формами с вполне развитым митозом , как красные водоросли , трудно понять с позиций прямой филиации , но он естественно вытекает из симбиотической теории .
Согласно теории прямой филиации и теории симбиоза , все организмы на Земле произошли от биохимически одно-типных бактериальных предков , а растениям и животным дали начало эукариотические микробы . Фотосинтезирующие бактерии дали начало водорослям , и в конце концов расте-ниям , а некоторые водоросли утратили свои пластиды и пре-вратились в предков грибов и животных . Теоретики прямой филиации согласны в том , что пластиды и другие органеллы эукариот , включая ядро , сформировались путем дифферен-циации внутри самих клеток и называют это ботаническим мифом .
Филогения типов растений
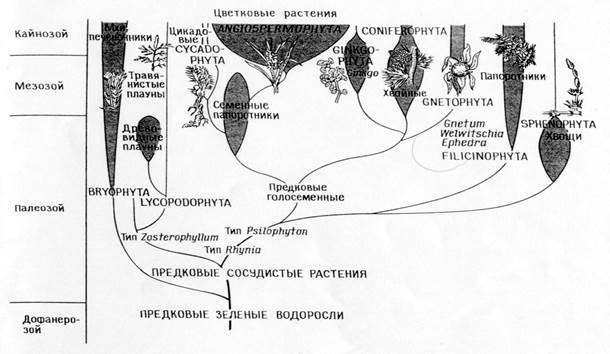
Сопоставление филогений , соответствующих теории прямой филиации (А)
и теории симбиоза (Б)
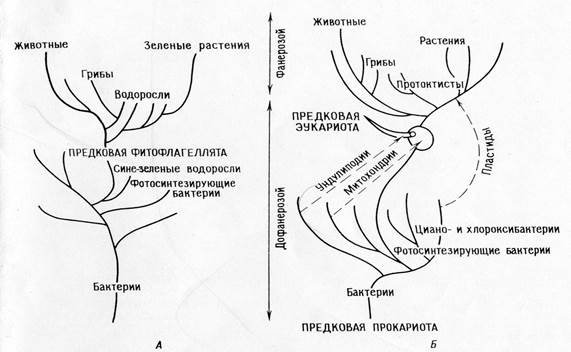
Различия между положениями теории прямой филиации и симбиотической теории :
Теория прямой филиации .
Главная дихотомия : животные – растения
Фотосинтезирующие эукариоты (водоросли и зеленые растения ) произошли от фотосинтезирующих прокариот ( сине-зеленых водорослей)
Растения и их фотосинтезируюшшие системы в докембрии эволюционировали монофилетически .
Животные и грибы произошли от автотрофов , утративщих пластиды .
Митохондрии дифференцировались внутриклеточно у фотосинтезирующих праводорослей .
У праводорослей внутриклеточно дифференцировались ундолиподии , митотический аппарат и все прочие эукариотические органеллы .
Все организмы произошли от предков – прокариот путем накопления одиночных мутаций .
СЛЕДСТВИЕ : согласно теории , должны существовать праводоросли ; трудно связать эту теорию с ископаемой летописью и построить соответствующую филогению .
Теория последовательных эндосимбиозов :
Главная дихотомия : прокариоты – эукариоты .
Фотосинтезирующие эукариоты (ядросодержащие водоросли и растения ) и нефотосинтезирующие эукариоты ( животные , грибы , простейшие ) произошли от общих гетеротрофных предков ( амебофлагеллят ).
Эволюция фотосинтеза в архее была монофилетической у бактерий ( включая циано- и хлорооксибактерий ), фотосин-тезирующие протисты возникли внезапно , когда их гетеро-трофные протисты – предки вступили в симбиоз с фото-синтезирующими прокариотами (протопластидами ).
Животные и большинство водорослей и грибов произошли непосредственно от гетеротрофных протистов .
Митохондрии уже были у гетеротрофных протистов ,когда последние в результате симбиоза приобрели пластиды . Промитохондрии были ранее приобретены симбиотическим путем .
Митоз выработался у гетеротрофных эукариотических протистов путем дифференциации ундолиподиальной системы .
Прокариоты произошли от ранних прокариотических предков путем ряда одиночных мутационных событий , эукариоты тоже произошли от эукариотических предков благодаря одиночным мутационным событиям , эукариотические же предки произошли от прокариот в результате серии симбиозов .
Следствия : теория объясняет биохимические пути и их избыточность у органелл . Она позволяет построить последо-вательную филогению , объясняющую “биологический разрыв “ между временами протерозоя и фанерозоя . Теория предска-зывает , что у органелл должны сохраняться остатки систем репликации .
Ботанический миф
Со времен Жюсье низшие растения называют таллофитами .а высшие - бриофитами и трахеофитами . Согласно ботаническому мифу ,общие предки фотосинтезирующих бактерий и сине-зеленых водорослей путем прямой филиации дали начало эукариотическим водорослям в том числе морским . Группы организмов , филогению которых в наибольшшей мере затра-гивает концепция прямой филиации , оказались в области ботаники - это бактерии , водоросли , грибы и растения .
Гипотетические организмы , которые могли бы быть связующим звеном между цианобактериями и зелеными водорослями , были названы родоначальными фито-монадами ( ancestral phytomonads ) или праводорослями . Некоторые авторы утверждали , что подходящими кандитами на роль праводорослей могут быть эукариоты Cyanidium и Cyanophora, сходные по своим пигментным системам с цианобактериями . У таллофитов не было обнаружено промежуточных форм между немитотическим состоянием цианобактерий , и вполне развитым типичным митозом зеленых водорослей и растений . Хотя грибы и цветковые растения полностью лишены ундулиподий , им свойственны митоз и мейоз , для которых необходимы веретена , состоящие из микротрубочек . Имеется общее согласие в том ,что все многоклеточные организмы ,обладающие микротрубочками ,происходят от эукариотических микроорганизмов с ундолиподиями , но никто не знает , от какой именно группы протистов . Подкреплением ботанического мифа служили древность и сложность фотосинтеза , а токже тот факт , что у многих организмов ( у бактерий , евгленовых , пара-зититических растений ) как в природе , так и в лаборатории , наблюдалась утрата фотосинтеза в результате мутаций . Поскольку фотосинтез - анаэробный процесс ,обеспечивающий в конечном итоге существование всех организмов , то он должен был развиться очень рано в истории жизни на нашей планете , еще до того как возник митоз , для которого небходим кис-лород . в фотосинтезе участвует множество пигментов , липидов и ферментов ,упорядоченно расположенных в очень сложных мембранах . При этом детали фотосинтетических механизмов у водорослей и растений , с одной стороны , и у цианобактерий , с другой - поразительно сходны . Все эти организмы освобождают газообразный кислород из воды , которая служит источником атомов водорода , используемых для восстановления СО2 до органических веществ клетки . Многие водоросли и простейщие не имеют ни митоза , ни генетической системы , сходной с таковой животных и растений . Инфузории имеют особую гене-тическую систему с ядрами 2 типов , причем клетки нередко многоядерные . Один тип яядер – макронуклеусы , а другой – намного меньшие микронуклеусы . Микронуклеусы физиологи-чески несущественны . однако они хранят копии генов , а при половом процессе - претерпевают мейоз . Физиологически необходимые макронуклеусы служат местом транскрипции РНК, и , таким образом , управляют физиологией клетки . Одна из трудностей при выяснении отношений между цианобактериями и эукариотическими водорослями связана с отсутствием данных о происхождении ундолиподий : не было найдено никаких промежуточных звеньев между цианобактериями , у которых ни-когда не бывает ундолиподий , и множеством водорослей , которые их имеют . Другая проблема – полное отсутствие полового процесса у цианобактерий : между бесполыми цианобактериями и водорослями , большинству которых свойственен половой процесс , не оказалось никаких промежуточных форм . Обнаружилось , что большинство водо-рослей имеют ундолиподии на какой – либо стадии жизненного цикла . Подвижные структуры у водорослей , простейших , жи-вотных и растений оказались одинаковыми : все они состоят из микротрубочек диаметром 0,024 мкм . Выяснилось ,что мито-тическое веретено даже у грибов , у которых нет ундолипдий ,состоит из белковых микротрубочек того же диаметра , что и в ундолиподиях . На протяжении полувека казалось чрезвычайно правдоподобным , что эукариотичкские водоросли произошли от цианобактерий . Этот ботанический миф привел к предположе-ниям , которые многие годы служили как стимулом для исследования , так и системой координат ,в которой про-изводилась оценка получаемых результатов . Утверждение , что связующие звенья между цианобактериями и эукариотическими водорослями вымерли , не оставив ископаемых следов и живых реликтов , становилось все менее убедительным по мере поступ-ления новых данных о живых микробах и об окаменелостях . Главные промежуточные звенья между фотосинтезирующими прокариотами и эукариотами не вымерли бесследно , как того требует ботанический миф ,- они никогда не существовали . Согласно теории последовательных эндосимбиозов , циано-бактерии – действительно родоначальники ,но только пластид , а не остальных частей эукариотической клетки , и уж во всяком случае не ее ядра . И все же в вопросе о происхождении ядра гипотеза прямой филиации и теория симбиоза совместимы . Мыслимые модификации крайнего варианта теории симбиоза включают , например ,идею о симбиотическом происхождении пластид , но не митохондрий , или о симбиотическом про-исхождении пластид и митохондрий , но не ундолиподий . Сторонники прямой филиации выдвинули также предположение о происхождении ядра путем симбиоза .
Решающие доказательства эндосимбиотического происхождения хлоропластов и митохондрий .
Наиболее существенные доказательства происхождения важней-ших органелл эукариотических клеток - хлоропластов и мито-хондрий из прокариот получены в последние годы при изуче-нии структуры рибосомальных РНК . Молекудярный биолог
К. Воз показал уже в 80-х годах , что структура РНК из рибосом хлоропластов растений почти идентична структуре РНК из некоторых цианобактерий . В то же время группа исследователей установила , что РНК рибосом , полученных из митохондрий разных эукариотов схожа с РНК некоторых бактерий , в частности бактерий рода Paracoccus . Набор дыхательных ферментов указанной бактерии очень похож на
“ дыхательный ансамбль “ митохондрий животных .
Эти данные достаточно ясно свидетельствуют в пользу большей вероятности эндосимбиотического происхождения эукариотических клеток . Явление эндосимбиоза и по сей день чрезвычайно широко распространено в природе . Известно , что на теле некоторых морских рыб часто поселяются светящиеся бактерии , которые не только сами получают большие преимущества для своей жизни , но и очень полезны для организмов хозяев . Все сказанное убедительно свидетельствует в пользу эндосимбиотического происхождения эукариотических клеток из гораздо меньщих по объему клеток прокариот . Однако главный вопрос заключается в установлении природы хозяйской клетки , ведь клетки эукариот обладают хорощо сформированным ядром , имеющим оболочку , а прокариоты такого ядра не имеют . Л. Маргелис , например , считает ,что роль хозяйских клеток , возможно , сыграли прокариоты – предшественники современных бактерий , относящиеся к микоплазмам - очень примитивно построенным прокариотам , практически лишенным клеточных стенок . В последнее время появились и широко обсуждаются работы японского ученого
Т. Ошимы . На основании тщательного изучения структуры рибосомных РНК многих про- и эукариот , в том числе и многих представителей до сих пор малоизученного надцарства архей , этот автор приходит к выводу , что хозяйской клеткой являлся один из предков современных архей . Следует сказать , что у представителей именно этого надцарства , в отличие от бактерий , имеется очень много общих с эукариотами биохи-мических и молекулярно – биологических особенностей . В по-следних работах Т. Ошима приводятся доводы в пользу того , что хозяйской клеткой при возникновении эукариот являлся предок архей , подобный одному из современных представителей этих прокариот - термоплазме . Каково будет окончательное решение проблемы установления природы хозяйской клетки , давшей начало эукариотической клетке в процессе множествен-ного эндосимбиоза , покажут дальнейшие исследования .
ВЫВОДЫ:
Наиболее популярной и обоснованной гипотезой происхождения эукариотических клеток является в настоящее время представле-
ние об их образовании путем многократно происходившего в процессе эволюции в течении первых 1.5 – 2 млрд лет существования на Земле живых клеток эндосимбиоза различных прокариот , относящихся к бактериям и археям . Решающие доказательства эндосимбиотического происхождения хлоро-пластов и митохондрий были получены ведущими биологами мира при изучении структуры рибосомальных РНК . Эти данные достаточно ясно свидетельствуют в пользу большой вероятности эндосимбиотического происхождения эукарио-тических клеток из гораздо меньших по объему (на несколько порядков ) клеток прокариот .
ЛИТЕРАТУРА
1. Соросовский образовательный журнал. Биология , Химия , Науки о земле , Физика , Математика . N 5 1998г
2 Маргелис Л. Роль симбиоза в эволюции клетки. Пер. с англ., М. МИР 1983г
3 Основы микробиологии . М . “Медицина “ 1992 .
4 Хахина Л.Н. Концепция А.С. Фаминцына о значении симбиоза в эволюции Л. ; Наука , 1981 , с . 165 – 181 .
5 Альбертс Б . Молекулярная биология клетки ; Пер. с англ . М . Мир 1986 т.1