| Скачать .docx | Скачать .pdf |
Курсовая работа: Имитационная модель динамики численности русского осетра
Федеральное агентство по рыболовству
ФГОУ ВПО «Астраханский государственный технический университет»
Институт рыбного хозяйства биологии и природопользования
Кафедра гидробиологии и общей экологии
КУРСОВАЯ РАБОТА
по дисциплине «Системная экология»
НА ТЕМУ:
ИМИТАЦИОННАЯ МОДЕЛЬ ДИНАМИКИ ЧИСЛЕННОСТИ ПОПУЛЯЦИИ РУССКОГО ОСЕТРА
Выполнила: ст. гр. ДБЭ-31
Саввинова Юлия
Проверил: к.б.н., Шипулин С.В.
Астрахань 2010
Содержание
ВВЕДЕНИЕ
ГЛАВА 1. ОБЩАЯ ХАРАКТЕРИСТИКА РУССКОГО ОСЕТРА
1.1 Ареал обитанияи систематика
1.2 Внешнее и строение
1.3 Миграция русского осетра
ГЛАВА 2. ВЛИЯНИЕ ФАКТОРОВ СРЕДЫ НА ДИНАМИКУ ЧИСЛЕННОСТИ ПОПУЛЯЦИИ РУССКОГО ОСЕТРА (Acipenser gueldenstaedtii Brandt)
2.1 Влияние абиотических факторов
2.1.1 Соленость воды
2.1.2 Влияние температуры
2.1.3 Влияние других факторов
2.2 Влияние биотических факторов
2.2.1 Межвидовая конкуренция
2.2.2 Паразитизм
2.2.3 Отношения «хищник - жертва»
2.3 Антропогенные факторы влияния
2.3.1 Промысел осетровых и его современное состояние
2.3.2 Загрязнение среды обитания
2.3.3 Влияние гидростроительства
2.3.4 Разведение и выращивание
2.3.5 Защита русского осетра
ГЛАВА 3. БЛОК-СХЕМЫ ИМИТАЦИОННОЙ МОДЕЛИ ДИНАМИКИ ЧИСЛЕННОСТИ ПОПУЛЯЦИИ РУССКОГО ОСЕТРА
ЗАКЛЮЧЕНИЕ
СПИСОК ЛИТЕРАТУРЫ
ПРИЛОЖЕНИЕ
ВВЕДЕНИЕ
Современные экологические исследования включаются в проблему, затрагивающую динамику развития сложных надорганизменных и внутриорганизменных процессов. В этих исследованиях большое значение имеет изучение механизмов воздействия факторов различной природы на организм осетра. Последствия такого влияния на русского осетра проявляются в различных формах в виде нарушения жизненных циклов, гаметогенеза, гибели на стадиях индивидуального развития, изменение популяционной структуры.
Изменение условий водной среды привело к снижению запасов русского осетра.
Оценка состояния запасов русского осетра определяется на основе изучения их биологии в различные периоды жизненного цикла. Изучается динамика и интенсивность нерестовых миграций, определяется численность особей, пропущенных на сохранившиеся естественные нерестилища Волги. По данным количественного учета скатывающихся личинок и молоди на стационарных пунктах наблюдений в реке оценивается пополнение в результате естественного нереста.
Полученные результаты свидетельствуют, что на фоне снижения численности русского осетра произошли значительные изменения в характере их распределения в Каспийском море. На традиционных местах концентраций осетровых плотность их скоплений в течение последних лет значительно снизилась, на отдельных участках - до нуля, что соответствует сокращению ареала распространения этого вида. Следовательно, наблюдаемые изменения в распределении осетра в Каспийском море вызваны не изменением локализации их концентраций, а снижением общей численности исследованных видов.
В естественных условиях обитания установлены случаи ухудшения морфо - функционального состояния организма, изменений в функционировании репродуктивной системы русского осетра.
Современное состояние запасов осетровых характеризуется резким сокращением численности на обследуемой акватории Каспийского моря, что нашло адекватное отражение в динамике их вылова. В популяциях рыб стала преобладать молодь, и это происходит не в результате увеличения пополнения запасов, а в результате чрезмерного изъятия промысловой части популяций.
Цель курсовой работы: Построить имитационную модель динамики численности русского осетра.
Задачи:
1. Изучить общую характеристику русского осетра.
2. Изучить экологические факторы, влияющие на имитационную модель численности русского осетра
3. Построить имитационную модель динамики численности русского осетра.
ГЛАВА 1. ОБЩАЯ ХАРАКТЕРИСТИКА РУССКОГО ОСЕТРА
ОСЁТР РУССКИЙ (Acipensergueldenstaedtii) Имеются проходная и жилая формы. Проходная форма имеет озимые и яровые расы. Яровые нерестятся в год захода в реки, озимые - на следующий год. Яровые формы медленнорастущие, озимые, наоборот, быстрорастущие (Берг Л. 1934).
Половое созревание волжского осетра наступает не ранее 10 лет, у самок не ранее 13 лет, куринского осетра — в 13— 14 лет, у самок — в 19—30 лет. Азовский осетр достигает половозрелости: самцы в 8—9 лет, самки в 10—14 лет; днепровский осетр созревает не ранее 11 лет. Наибольший известный возраст — 48 лет, наибольшая длина — 230 см и вес (изредка) — до 80 кг и даже 120 кг. Средний промысловый вес волжского осетра 12—16 кг. В 1960 г. средний вес волжского осетра был 14,5 кг, в 1962 г. средний вес самцов ходового осетра на Волге был 11,7 кг, самок — 21,3 кг. Средний промысловый вес куринского осетра 22—24 кг, азовского — около 15 кг. Плодовитость русского осетра составляет от 84 до 837 тыс. икринок, в среднем 250—350 тыс. икринок. В Каспийском море осетр питается в раннем возрасте беспозвоночными (гаммариды, мизиды, и др.), с возрастом переходит на питание моллюсками и рыбой (бычки, сельди, кильки). Осетр в природе образует помеси с белугой, севрюгой, шипом, стерлядью. Как уже говорилось выше, на Волге, в районе Саратова, путем искусственного оплодотворения получены жизнестойкие гибриды: осетр X стерлядь, стерлядь X осетр, осетр Х белуга и белуга Х осетр (Ходоревская Р.П., с соав. 1997).
1.1 Ареал обитания и систематика
Русский осетр населяет бассейны Каспийского, Черного и Азовского морей. Имеются проходная и жилая формы. Проходная форма имеет озимые и яровые расы. В Волге, Каме и, по-видимому, Урале есть жилые (постоянно живущие в реке) формы. В Каспии наиболее многочисленно волжское стадо. В Азово-Черноморском бассейне осетр образует стада: черноморско-кавказское (рионское), черноморско-украинское (днепровское) и азовское. Для икрометания из Каспия входит в Волгу, меньше в Урал, в очень незначительном количестве заходит в Терек, Сулак, Самур. По иранскому побережью осетр входит в Сефидруд, изредка в Горган, Баболь и другие реки. Из Черного моря осетр входит в Дунай, в Днепр, очень немного в Риони, Мзымту, Псоу и другие реки. Из Азовского моря на икрометание входит в Дон и единичными экземплярами в Кубань; в последние годы ход осетра в Кубань (как и белуги) увеличивается (Никольский Г.В. 1974).
Основная нерестовая река — Волга, по которой ранее осетр поднимался до Ржева, а также в многочисленных ее притоках — Шексне, Оке, Ветлуге, Каме, Вишере и др. По Уралу поднимался до Оренбурга. Из Каспия заходит в небольшом количестве также в Терек, Сулак и Самур. Из Черного моря осетр входит в Дунай и Днепр, очень мало в Риони, Мзымту, Псоу и другие реки; по Днепру поднимается до Могилева и изредка до Дорогобужа (Ходоревская Р.П., с соав. 1997).
ЦарствоRegnum: Animalia
Тип Phylum: Chordata
Подтип Subphylum: Vertebrata
Инфротип Infraphylum: Gnathostomata
Надкласс Superclassis: Osteichthyes
Класс Classis: Actinopterygii
Подкласс Subclassis: Chondrostei
Отряд Ordo: Acipenseriformes
Семейство Familia: Acipenseridae
Подсемейство Subfamilia: Acipenserinae
Вид Species: Acipenser gueldenstaedtii
1.2 Внешнее строение
Внешние признаки: 10-13 спинных жучек с острыми, загнутыми назад концами (у старых особей затуплены), 21-50 мелких боковых жучек, удаленных друг от друга настолько, что между ними видна боковая линия, 8-10 брюшных жучек. Сильное веретенообразное тело выше, чем у остальных осетровых, рыло широкое и короткое. Перед маленьким щелевидным ртом находятся короткие круглые усики без бахромок; отогнутые назад, они не достигают края рта. Нижняя губа, как правило, посередине прервана. Окраска: спина от серо-голубого и пепельно-серого цвета до оливково-серого, бока светлее, брюшко от беловатого до грязно-желтого. Костные щитки цвета слоновой кости. Длина: обычно 130-250 см (20-30 кг), макс. до 400 см (до 160 кг) (рис.1.) (Богатова И.Б. 1980).
 |
Рис.1. Внешний вид Русского осетра.
Отделы тела осетра - голова, туловище, хвост - плавно переходят друг в друга, обеспечивая обтекаемость. Выделяемая многочисленными кожными железами слизь уменьшает трение при движении рыбы. Парные плавники грудные и брюшные - поддерживают нормальное положение тела спиной вверх, служат рулями поворота.
Органы дыхания – жабры - расположены на верхней стороне четырех жаберных дуг в виде ярко-красных лепестков.
Кровеносная система осетров заткнута. Сердце двухкамерное, состоящее из предсердия и желудочка.
Орган слуха и равновесия представлен только внутренним ухом, расположенным по бокам задней части черепа.
Размножение. Парные половые железы — яичники и семенники — имеют выводящие протоки. Оплодотворение наружное и происходит в воде (Вишнякова М.А., Брудастова Р.И.1989).
1.3 Миграция русского осетра
Главная масса осетров держится в самом море, хотя и в пресноводной или малосоленой его части. Отсюда, начиная с апреля, он начинает входить в реки для икрометания. Идет он обыкновенно малыми косяками и придерживается, как и в другое время, самых глубоких и быстрых мест реки; бег его тише севрюги, но быстрее, чем у шипа. В Днепре в мае начинается уже обратный ход его, но в Урале и Волге он остается в реке значительно дольше, а в Енисее возвращается вниз к 25 августа. По свидетельству Северцева, осетры, которые здесь малочисленнее белуг, начинают входить в Урал с середины апреля. Ход их зависит, как и у всех проходных рыб, от попутного ветра: при противном ветре они также толпятся у мелей перед устьями и ждут моряны, чтобы вместе с волной войти в реку. Число их особенно увеличивается с половины мая, но осетры, входящие в реку в конце этого месяца и в июне, уже не мечут здесь икры, а остаются зимовать на ятовях и нерестятся уже в следующую весну.
Ход рыб представляется в следующем виде: молодые осетрики (костерята), скатившись в море, через несколько (4- 5) лет достигают там половой зрелости, входят весной в реку, мечут в низовьях, а затем в скором времени скатываются обратно в море; на следующий год они также входят в реку, но уже не весной, а летом, остаются в нижнем течении, кормятся там и зимуют в глубоких речных ямах; на третий год поднимаются из ям и мечут в более верхних течениях реки. Отсюда, конечно, следует заключить, что все или же большая часть осетров, мечущих икру в Каме, в верхнем течении Волги, пришли сюда не из моря. Но почему здесь осетры мечут ранее, нежели в низовьях,- это объяснить довольно трудно и остается предположить, что икра у осетров, зимующих в реке, развивается быстрее, нежели у таких, которые входят для нереста из моря. В Волге, начиная от Рыбинска до Самары, осетр нерестится почти одновременно со стерлядью или немного позже- в первой половине мая. Это подтверждает как проф. Кесслер, так и акад. Овсянников, которому именно в это время удалось получить зрелые молоки осетра и даже оплодотворить ими икринки стерляди. Между тем, по исследованиям академика Бэра, в низовьях Волги, около Сарепты и Царицына, а также в реке Куре осетр мечет икру не раньше конца июня и даже преимущественно в июле. То же самое замечено О. А. Гриммом и под Саратовом. Однако в Урале и Сефидруде вся красная рыба, по Бэру же, начинает нереститься в исходе апреля и оканчивает в начале июня, что противоречит высказанному нами предположению о влиянии пресной воды на развитие икры красной рыбы.
Нерест происходит, как у других осетровых, на глубоких и быстрых местах реки с каменистым или хрящеватым дном. Он весьма непродолжителен — 3-4 дня. Созревание и метание икры происходит весьма быстро, и вся икра делается совершенно зрелой почти одновременно у каждой особи и одновременно же вся выметывается. Это доказывается тем, что как в начале, так и в конце периода метания икры встречаются осетры с вытекающими молоками и икрой.
Молодые осетры довольно долгое время живут в тех местностях, где выклюнулись из яичек, а затем скатываются в море, где и пребывают до достижения ими половой зрелости - пяти лет, даже более. Сколько времени осетры остаются в реке - достоверно неизвестно; по словам академика Бэра, они уходят по достижении одного года, и двухлетки никогда в реке не встречаются Двухлетние осетры держатся вместе со стерлядями.
Главная масса молодых осетров уходит в море, в ту же осень или в весеннее половодье, т. е. не достигнув годового возраста (Ходоревская Р.П., с соав. 1997).
ГЛАВА 2. ВЛИЯНИЕ ФАКТОРОВ СРЕДЫ НА ДИНАМИКУ ЧИСЛЕННОСТИ ПОПУЛЯЦИИ РУССКОГО ОСЕТРА (Acipenser gueldenstaedtii Brandt)
Русский осетр, как и другие виды, подвергаются воздействию различных факторов: биотических, абиотических и антропогенных.
2.1 Влияние абиотических факторов
К абиотическим факторам относятся все факторы неживой природы. Они в той или иной степени влияют на рост, развитие и жизнедеятельность осетра. Основными факторами являются температур, соленость и свет.
2.1.1 Соленость воды
Установлено, что в процессе развития осетровых, происходит быстрое повышение солеустойчивости, но есть особенности. Так, типично проходные виды (севрюга, русский осетр), достигнув массы 2-4 грамма, способны выдерживать соленость до 12‰. Белуга же подобную соленость выдерживает при массе 4-8 граммов. Пресноводные осетровые, такие как сибирский осетр, большой амурский лопатонос, веслонос негативно реагируют на повышение солености воды. Так, личинки веслоноса массой 0,1 г не выдерживают солености выше 5‰ при температуре воды 20°C и 7,5‰ при 13-14°C. Мальки даже весом 1 г гибнут в воде с соленостью 10‰.
У личинок осетра при переходе на активное питание соленость 2-3‰ положительно влияет на прирост. У молоди в возрасте 15-20 дней после начала питания наиболее быстрый рост наблюдался уже при солености 6‰. Среднесуточный привес составил 15%, тогда как в пресной воде - 10%.
В 30-дневных опытах с постепенным возрастанием солености воды с 3 до 8‰ личинки и мальки (43 мг) русского осетра на 20% превысили массу контрольных особей в пресной воде.
Для каждого вида и возраста рыб есть свой оптимальный уровень солености воды. Так, активный рост русского осетра и севрюги в течение первых 10 дней активного питания улучшался при увеличении солености воды до 35‰, у белуги при 2‰, у шипа — в пресной воде. Более крупные белужата весом 13-16 г уже лучше росли в воде соленостью 7‰, а шип массой 2 г тоже дал лучший прирост при 7‰.
И практика, и эксперименты подтверждают, что для молоди осетровых, выращиваемой в слабосоленой воде, характерны повышенный аппетит и более быстрый рост (Денисов А.И. 1978).
2.1.2 Влияние температуры
Исследования скопления осетровых в море в пространстве и глубине показали, что прямо или косвенно их нахождение связано с температурой воды. В большинстве случаев осетровых наблюдали на глубине 6-8, 18-24 и 40-55 метров. Молодь русского осетра в наибольшем количестве встречали при температуре 25-26°C, меньше — при 23-25°C и 26-28°C.
Разные виды осетровых, имеют свой оптимум температуры, который с возрастом меняется. Опытным путем установлено, что, например, оптимальным температурным режимом для развития икры осетра осеннего хода является 20-23°C. Личинки в возрасте 1-2 суток не способны выбирать оптимум температуры. Но уже в возрасте 3-10 суток выбирают температурную зону 20-22°C, 10-20 суток — 18,5-20,5°C, 20-35 дней — 16-18,5°C, 35-45 суток - 14,5-16°C, а 45-60 суток — 12-14,5°C.
При выращивании личинок в таких температурных условиях рост, интенсивность питания и выживаемость были самыми высокими. При повышении температуры до 27°C гибели осетровых не наблюдали. Оптимальными температурами для развития осетровых рыб является диапазон 10-22°C. Температурные оптимумы у ранней молоди различных видов осетровых находятся в следующих пределах: для белуги 8-15°C, севрюги 16-22°C. Осетр по термоустойчивости ранних стадий занимают промежуточное состояние между белугой и севрюгой. Диапазон чувствительности к понижению температуры ранних стадий у белуги 4-7°C, у севрюги 6-10°C.
В первые дни активного питания личинок температура воды, при которой скорость роста наибольшая, ниже всех у белуги — 17,5-19,5°C и возрастает в ряду стерлядь — шип — русский осетр — севрюга. У севрюги максимальная скорость роста наблюдается при 24-25°C. Характерно, что температурная последовательность соответствует временной, при которой эти виды идут на нерест.
Оптимальная температура для развивающейся молоди осетра русского волжского — 14-18°C, а куринского — 12-18°C.
Устойчивость к температурным колебаниям зависит от солености воды. Русский осетр перевод из воды с температурой 20°C в воду с температурой 30°C переносит хорошо. Но в соленой воде до 16‰ такие скачки температур вызывают гипернатриемию и гибель рыб.
Для взрослых осетровых в стадии нереста оптимальный спектр температуры воды является 7-26°C.
Популяцию русского осетра реки Лены необходимо рассматривать как самую холодолюбивую из всех осетровых. Ленский осетр обладает способностью питаться и расти в зимнее время при низкой температуре воды. Его рост, особенно линейный, к зиме не замедляется столь значительно, как у других осетровых.
Как бы ни была изумительна способность приспосабливаться к холодным водам, наиболее высокий темп роста осетровых происходит на источниках с постоянной теплой водой, где и продолжительность активного питания существенно увеличивается. Однако необходимо знать и противоположную, негативную сторону. В исследованиях определения зависимости среднесуточного привеса массы при различных рационах питания и эффективности использования пищи на рост личинками веслоноса (масса 0,07-0,1г) от температуры установлено следующее. Наиболее высокие показатели получены при температуре воды 18°C. При рационе, составляющем 10% массы тела, особи, содержащиеся при температуре выше 18°C, погибли. При концентрации, обеспечивающей рацион 40% массы тела, при температуре 23°C теряли в весе, а при концентрации пищи, равной 80% массы, тела рыбы росли даже при 28°C. Особенность состоит в том, что с увеличением температуры воды с 18 до 28°C скорость прироста веса рыб, рацион которых составлял 80% массы их тела, уменьшалась в связи с тем, что увеличивался энергетический обмен, и снижалась эффективность использования пищи на рост.
Высокие температуры воды оказывают на рыб негативное, а порой и губительное действие, которое объясняется не только тепловым повреждением клеток, но и ухудшением условий дыхания. С повышением температуры воды потребность рыб в кислороде увеличивается на максимуме почти в два раза, при этом повышается и граница кислородного порога.
Следить за температурой воды в водоемах или искусственных сооружениях необходимо постоянно, особенно при переходе оптимальных температур в сторону повышения или понижения. Температура ниже или выше оптимума снижает продуктивность рыб и в определенных пределах может привести к гибели организмов (Привезенцев Ю.А. 1986).
2.1.3 Влияние других факторов
Влияние освещенности:
Изучено влияние постоянной и переменной освещенности на мальков русского осетра. При изменении освещенности от 30 до 800 лк повышалась скорость роста мальков, увеличивались интенсивность дыхания, суммарное потребление корма и кислорода, но снижались кормовой коэффициент и расход кислорода на прирост. При переменной освещенности с определенной амплитудой и периодичностью увеличивались скорость роста, снижался расход кислорода на прирост единицы массы и улучшалось усвоение пищи. При переменном режиме освещенности, оптимальном для роста мальков, отмечается моноцитопения и лимфофилия. Количество общих липидов и белка в мышцах осетра достоверно повышалось, а обводненность снижалась при переменном режиме освещенности (Вишнякова М.А., Брудастова Р.И. 1985).
Прозрачность воды:
По сравнению с воздухом вода гораздо менее;-прозрачна и попадающий в нее свет довольно быстро поглощается и рассеивается. При прохождении через толщу воды меняется спектральный состав света, что существенно влияет на условия фотосинтеза и отражается на поведении русского осетра.
Прозрачность воды является одним из основных критериев, позволяющих судить о состоянии водоема. Она зависит от количества взвешенных частиц, содержания растворенных веществ и концентрации фито- и зоопланктона. Влияет на прозрачность и цвет воды. Чем ближе цвет воды к голубому, тем она прозрачнее, а чем желтее, тем прозрачность ее меньше.
Важным фактором, определяющим прозрачность воды в непроточных водоемах, являются биологические процессы. Прозрачность воды тесно связана с биомассой и продукцией планктона. Чем лучше развит планктон, тем меньше прозрачность воды. Таким образом, прозрачность воды может характеризовать уровень развития жизни в водоеме. Она имеет большое значение как показатель распределения света (лучистой энергии) в толще воды, от которого зависят в первую очередь фотосинтез и кислородный режим водной среды (Бурмаков Г.Т., Моисеев Н.Н. 1981).
Кислород:
Мощным источником обогащения воды молекулярным кислородом является фотосинтез водных растений, интенсивность которого зависит от температуры и освещения. Фотосинтез происходит главным образом в поверхностных слоях воды, хорошо освещенных и прогретых.
У осетра отмечается четкая видовая специфичность как в отношении минимального количества кислорода, растворенного в воде, при котором может жить рыба, так и в отношении интенсивности потребления кислорода в процессе дыхания. При увеличении температуры воды пороговое напряжение кислорода возрастает Влияние кислородных условий на эмбриогенез это связано в первую очередь с изменением скорости развития и роста. Так, с увеличением содержания кислорода в определенном диапазоне концентраций происходит ускорение эмбриогенеза. Дальнейшее увеличение содержания кислорода приводит к замедлению развития зародышей и углублению образующихся аномалий. Известно, что избыточная концентрация кислорода может быть даже летальной.
От концентрации кислорода в воде зависит их жизнедеятельность. При уменьшении его ниже определенных границ падает интенсивность питания и использования пищи на рост, в результате чего замедляется рост. В условиях интенсивного рыбоводного хозяйства снижение скорости роста наступает при уменьшении содержания кислорода от 40 до 65 %.
При недостаточном содержании кислорода в воде снижается устойчивость к неблагоприятным факторам внешней среды, в том числе к промышленным и бытовым загрязнениям. Низкое содержание кислорода обусловливает неблагоприятные зоогигиенические условия в водоеме, в результате чего создаются предпосылки к накоплению органических веществ и размножению сапрофитной микрофлоры, которая может отрицательно воздействовать на осетра. Длительное пребывание в воде с недостаточным содержанием кислорода понижает активность , резко снижает устойчивость к возбудителям болезней(Данилов Ю.М. 1980).
Углекислый газ:
Отрицательное влияние высокой концентрации углекислоты на жизнедеятельность осетра заключается в том, что находясь в угнетенном состоянии, хуже используют кислород, растворенный в воде. При этом значение имеет не просто абсолютное содержание в воде кислорода и углекислоты (диоксида углерода), а их соотношение. При низком содержании кислорода и неблагоприятном соотношении кислорода и углекислого газа осетр значительно хуже использует корм. Критическая концентрация углекислого газа для взрослой особи280-300, для молоди -200, для личинки -160 (Гербильский. Васильева Л.М. 2000).
Вертикальное распределении:
В основном встречаются на глубинах 2-130 м, зимой - 10-40 м, а весной - 10-30 М. Летом русский осетр мигрирует в наиболее мелководные районы моря с глубинами менее 20 м, а осенью перемещается на глубине до 100 м (Легеза, 1972).
Влияние колебания уровня моря:
В годы падения уровня Каспийского моря, мальки осетров, которые спустились вниз по течению рек Волга и Урал, мигрировали непосредственно с западного шельфа Среднего Каспия, не задерживаясь в северной части моря. В многоводные годы осетры более широко распространены в Северном Каспии, чем в маловодные годы(Исаев А.И. 1988).
2.2 Влияние биотических факторов
Помимо абиотических факторов, на русского осетра, также влияют биотические факторы (живой природы). К ним относятся такие факторы, как конкуренция, питание (отношения хищник – жертва), влияние паразитов, и различные другие факторы.
2.2.1 Межвидовая конкуренция
Межвидовая конкуренция во время кормления на море преобладает между русским осетром и теми видами рыб, с которыми совпадает спектр продуктов - бычки, севрюга, белуга. В последние годы, увеличилось совпадение в пищевом спектре осетра и карповых (вобла, лещ). Конкуренция была выявлена в отношении таких продуктов питания, как килька и бычки (Карпевич Л.Ф. 1975).
2.2.2 Паразитизм
Паразиты редко являются проблемой для осетров. Но большое количество внешних паразитов могут повредить рыбе, и особенно дыхательным путям. Могут быть обнаружены такие простейшие как Trichodina, Trichodonella and Ichthyophthirius multifiliis (вызывает образование белых пятен) и жгутиковые, такие как Ichthyobodo (= Costia). В меньшей степени может быть обнаружена Ambiphrya. Все эти паразиты являются внешними простейшими паразитами. Наиболее чувствительная естественно мелкая рыба, малёк и личинка. Жгутиковая Hexamita может быть обнаружена в кишечнике малька осетра. Такие жгутиковые присутствуют в большом количестве у многих видов рыб. Эти паразиты вызывают тяжёлое заболевание жабр (Амёбное жаберное заболевание или АЖЗ) аналогичное тому, которое можно наблюдать у других видов рыб. Из-за АЖЗ наблюдалась сильная гиперплазия и спайка жаберных пластин, что вызвало высокую смертность(Козлов В.И., Абрамович Л.С. 1986).
У осетров встречаются, как правило, общие виды паразитов из числа тех, что вызывают у них заболевания или же могут повлиять на их пищевое использование. По этой причине ниже приведена общая паразитологическая характеристика рыб рода русский осетр.
Основные болезни и паразиты
Поверхность тела:
1. На поверхности тела осетров, вылавливаемых в Каспийском море, иногда встречаются некрупные пиявки – каспиобделла каспийская (Caspiobdellacaspica).
2. На теле русского осетров у основания плавников, в хвостовой части и между костными жучками поселяется копепода – псевдотрахелиаст звeздчатый, или каспийско-черноморский (Pseudotracheliastesstellatus). Тело рачка состоит из 3 отделов: головогруди, максиллы II (так называемые “руки”) и туловища. Головогрудь маленькая; туловище уплощeнное, длинное, крупное, на его спинной стороне видны два ряда маленьких углублений; яйцевые мешки длинные, тонкие. Длина тела 1,7 – 2 см, “рук” до 1,2 см, яйцевых мешков до 2,7.
Рачки разрушают покровные ткани рыбы и глубоко проникают в еe мускулатуру, образуя раны. Вокруг прикрепительного аппарата рачка формируется капсула, заполненная гноем. Очень часто при вылове рыбы паразит отрывается, а место его прикрепления можно определить по глубоким (2 – 3 см), гноящимся язвам. Заражeнность осетров псевдотрахелистом может достигать 100 %, а количество рачков на одной рыбе – 50 экз. Заражeнность севрюг также довольно высока – 75 %, по 1 – 30 экз. на рыбе.
Псевдотрахелиаст найден у осетровых в бассейнах Чeрного, Азовского и Каспийского морей. В Азовском море паразит заметно чаще встречается летом, в сравнении с остальными сезонами года.
Внутренние органы:
Икру поражает микроспоридия рода плейстофор – Pleistophorasulci (син.: Cocconemasulci). Заражeнные икринки белого цвета, крупнее здоровых. В центре икринок находятся панспоробласты со зрелыми спорами паразита, а по периферии – споронты и панспоробласты с молодыми спорами.
Заболевания встречается повсеместно, где обитает осетр.
Икра бывает инвазирована полиподиумом гидроподобным. Икра заражается в период образования в ней желтка. По мере роста икринки личинка паразита разрастается, преобразуется в так называемый столон, на котором образуются почки, зачатки будущих самостоятельных особей. Заражённые икринки выходят в воду вместе со здоровыми. В воде оболочка икринки разрывается, столон выходит наружу и распадается на составляющие его почки, каждая из которых становится самостоятельным полипом (Михеев В.П. 1982).
2.2.3 Отношения «хищник – жертва»
В основном русский осетр питается придонными организмами. В поисках пищи осетр непрерывно движется в нижней части толщи воды. Вкусовые рецепторы расположены на усиках и губах. Во время охоты осетр делает резкое движение захвата и всасывает пищу с выдвиганием устного аппарата.
В Волге личинки осетра кормятся зоопланктоном - дафнии, Bosmina, циклопы. В реке Урал потребляют личинок донных организмов, а также: олигохет, полихет, Corophiidae, гаммарид, Mysidae, личинок и куколок хирономид. В ходе вниз мальки охотятся на нижних и придонных организмов: гаммарид, Mysidae, Corophiidae, олигохет и Ampharetidae. На море, диета несовершеннолетних осетров (41-80 см TL) содержит меньше ракообразных и Nereis, потребление крабов и рыбы небольшой. Как осетр вырастает, он переходит на питание, в основном, моллюсками, преобладающие виды - Абра Овата, представитель фауны Средиземноморья внесен в Каспийское море. Взрослый осетр поедает бентофагов и моллюска-людоеда. Его спектр питания изменяется в зависимости от поставок продовольствия и сезона (Детлаф, Таас Гинзбург, О. И. Shmalgauzen 1981) .
2.3 Антропогенные факторы влияния
Значительная часть прибрежных и морских экосистем России подвержена антропогенным нарушениям.
Антропогенные факторы, действующие на морские экосистемы разнообразны и, как правило, имеют комплексный характер и множественные экосистемные последствия.
2.3.1 Промысел осетровых, и его современное состояние
Хозяйственное значение. Ценнейшая промысловая рыба, у которой ведущее место принадлежит каспийской популяции. В период с конца 40-х до начала 60-х годов на Каспии ежегодно вылавливалось от 8,5 до 17,9 тыс. т русского осетра. Рекордный улов был достигнут в 1903 г. и составил 39,2 тыс. т.(таб. 1.). После постройки Волгоградской ГЭС его численность и уловы стали постепенно падать. В 1985 г. общая численность нагуливающегося в Каспийском море русского осетра (от годовика и старше) составила 59,1 млн экз., в 1994 г. она снизилась почти в три раза — до 21,2 млн экз. Плотина Волгоградской ГЭС отрезала 80% нерестилищ русского осетра и, хотя этот вид является основным объектом искусственного разведения, результаты его пока не достигли желаемого уровня, и в пополнении запасов все еще решающую роль играют оставшиеся ниже плотины нерестовые площади. После распада СССР численность русского осетра на Каспии катастрофически снижается, возобновлен морской лов, резко усилилось браконьерство и т.д. (Милн П. 1978).
Таблица 1. Численность русского осетра на акватории Каспийского моря
| годы | Численность русского осетра (млн. экз.) |
| 1968 - 1970 | 93,7 |
| 1972 – 1978 | 69,2 |
| 1983 – 1988 | 46,6 |
| 1991 – 1999 | 19,6 |
| 2001 - 2005 | 10,5 |
2.3.2 Загрязнение среды обитания
Загрязнение представляет опасность в первую очередь для мелководных полузамкнутых морских бассейнов, расположенных в зоне интенсивной индустриализации Северного Каспия. Здесь во многих районах концентрация загрязнителей в среде и биоте во много раз превышает ПДК. Огромное количество загрязнений поступает в моря с водами рек
Особенно опасно то, что в морских водоемах загрязнители, включая радиоактивные вещества, аккумулируются в донных осадках(Мельников В.Н. 1983).
Хроническое Северного Каспия явилось одной из причин изменений нерестового поведения, интенсивности и сроков подхода зрелых рыб к береговой зоне моря, нарушениями в репродуктивной системе рыб, что с течением времени (через 50-100 лет) может привести к полной деградации генофонда популяций. Ведущая роль среди токсикантов в развитии этих негативных изменений принадлежит свинцу, а также ртути и группе метаболитов ДДТ.
Эвтрофикация эстуариев и шельфовых мелководий. Эти процессы явились результатом резкого увеличения в последние годы стоков бытовых и промышленных отходов с высокой концентрацией биогенных элементов, а также смыва удобрений с сельскохозяйственных полей. Повышение концентрации биогенов в воде вызывает бурные вспышки развития микроводорослей. Эвтрофирование проявляется в "цветении" воды из-за интенсивного развития фитопланктона в пик вегетации и создает дефицит кислорода в воде. Результат - уменьшению численности русского осетра и их пищевых объектов, сокращение видового разнообразия.
Эвтрофикация является одним из факторов, приводящих к росту уязвимости экосистем и инвазиям чужеродных видов (Козлов В.И. 1977).
Судоходство. К числу существенных негативных факторов следует отнести судоходство. На мелководных участках с глубиной 5-10 м морские суда, имеющие осадку 3-5 м, нарушают естественную среду обитания организмов. С судоходством связаны дноуглубительные работы, шумовые эффекты, явление кавитации, взмучивание илистых грунтов, разрушение лесозащитных полос вдоль берегов, загрязнение акватории нефтепродуктами. Только в одном Азовском море через прорытые по мелководьям судоходные каналы уже сейчас ежегодно проходят 7 тыс. судов. И этот поток будет возрастать.
Крайне опасное побочное действие судоходства на морские экосистемы – расселение экзотической фауны. С балластными водами или на днищах судов морские организмы распространяются далеко за пределы своего естественного ареала. Многие виды-вселенцы, обладая широкой экологической пластичностью и высокой скоростью размножения, формируют в новых водоемах многочисленные популяции и существенно изменяют структуру биоценозов.
Распространение чужеродных видов. Эта проблема характерна для многих морей Мирового океана. Вселенцы внедряются двумя путями: случайно или искусственно. Многие виды - вселенцы обладая широкой экологической пластичностью и высокой скоростью воспроизводства, формируют в новых водоемах многочисленные популяции и существенно изменяют видовую и пищевую структуру экосистем. Вторжение чужеродных видов часто приводит к колоссальным вспышкам их численности, подавлению аборигенных видов, разрушительным трансформациям местных экосистем и огромным убыткам.
Таким образом, добавление всего одного нового элемента привело к катастрофическим трансформациям морских экосистем всего Азово-Черноморского региона, что в свою очередь подорвало кормовую базу русского осетра (Никоноров А.М с соав. 2000).
Нарушение структуры морских экосистем в результате антропогенных воздействий.
Загрязнения:
1. нефтепродуктами и буровыми растворами от добывающих предприятий, а также в результате аварий трубопроводов (юго-восточная часть Баренцева моря, северная и средняя части Каспийского моря, северо-восточный шельф Сахалина);
2. выбросами морского транспорта и морских портов;
3. отходами промышленных предприятий и бытовыми стоками: тяжелыми металлами, фенолами, поверхностно-активными веществами и другими загрязнителями;
4. отходами сельского хозяйства: соединениями фосфора, азота, пестицидами и др.;
5. радиоактивными и токсическими веществами в результате военных испытаний, функционирования военных полигонов и баз, захоронения отходов;
6. строительные работы и добыча полезных ископаемых в прибрежной зоне;
7. инвазии чужеродных видов;
2.3.3 Влияние гидростроительства
Изменение экологических условий в бассейне вследствие взаимодействия природно-климатических факторов и хозяйственной деятельностью человека обусловливает колебания численности русского осетра. Особенно значительное воздействие на экологию бассейна и воспроизводство рыбных запасов оказало гидростроительство. Нарушение естественного гидрологического режима р. Волги в результате гидростроительства и эксплуатации ГЭС привело к ежегодной потере более 180 тыс. т ценных промысловых рыб (осетровые, сельди, вобла, сазан и др.). Значительно утратили свое рыбохозяйственное значение реки Кура, Терек, Сулак, Самур. Ухудшились условия размножения рыб на Волге и Урале: площади нерестилищ осетровых на р. Волге сократились с 3600 до 430 га (Иванов, 2001).
В результате зарегулирования стока Волги каскадом плотин нерест осетровых в настоящее время возможен только на участке, расположенном ниже плотины Волгоградской ГЭС (Дубинин, 2002). В нижнем бьефе Волгоградского гидроузла наблюдаются резкие суточные и недельные колебания уровня воды и скорости течения, чего не наблюдалось до зарегулирования стока реки Волги. Это приводит к обсыханию нерестилищ, создает на них неустойчивый скоростной и температурный режимы, что сокращает нерестовые площади, вызывает гибель отложенной икры (Королева В.А. 1985).
2.3.4 Разведение и выращивание
Формирование промысловых запасов осетра происходит в настоящее время за счет естественного и искусственного воспроизводства (Баранникова, 2002). Наибольшее количество молоди осетра выпускают волжские заводы. В период летнего нагула осетр придерживается глубины 10-25 м. Осенью и зимой он откочевывает на большую глубину, иногда до 100 и более метров. Зимой наибольшее значение для нагула приобретают районы западного побережья Среднего Каспия и восточное побережье Южного Каспия. В пределах ареала осетр совершает сезонные перемещения, чаще всего связанные с температурным режимом воды и распределением кормовых организмов. По характеру питания осетр является бенто-фагом-моллюскоедом.
В популяции осетра в последние годы увеличилась доля молоди и особей непромысловых размеров, снизились показатели абсолютной длины, массы и возраста рыб, уменьшился процент самок, снизились показатели относительной и абсолютной численности (Иванов, 2000; Левин, 2001). Все это свидетельствует об ухудшении условий формирования общего запаса осетра в море, что подтверждается снижением численности, и биомассы нерестовой части популяции волжского осетра.
Существенную роль в воспроизводстве осетровых пород рыб играет их заводское разведение. В целях компенсации ущерба, нанесенного гидростроительством, в СССР с 1954 года было построено 13 осетровых рыбоводных заводов. В 1980-1990е годы ежегодный выпуск 2-3граммовой молоди достигал более 100 млн. экземпляров. Около половины молоди вывозилось с заводов, на специальных судах, в море к местам нагула во избежание истребления ее хищниками в процессе ската по реке. Коэффициент промвозврата в разные годы менялся, но чаще всего составлял для русского осетра около 1%. В настоящее время объемы промышленного воспроизводства сократились и являются значительно меньшими, чем величина изъятия из популяции.
Более 50% запасов каспийских осетровых формируется за счет выпуска молоди рыбоводными заводами, преимущественно российскими. По состоянию кормовой базы выпуск молоди осетровых в море следует существенно увеличить и довести до 150 млн. штук в год с преобладанием молоди осетра. При этом важной задачей является повышение жизнестойкости молоди и сохранение ее на местах нагула в море, что требует принятия мер по предотвращению браконьерства и углеводородного загрязнения среды обитания. Для транспортировки заводской молоди осетровых в море, на судостроительных заводах России, создаются новые живорыбные суда. Осуществляется техническая модернизация предприятий с учетом новых элементов биотехники разведения.
Перевозка молоди осетра:
В процессе выращивания осетровых возникает проблема перевозки молоди. Первая задача заключается в создании оптимальных условий при перевозке и доставке живой рыбы к месту назначения. Молодь массой 3-5г можно доставлять в фермерское хозяйство 1втоцистернами. При перевозке на большие расстояния (более 6ч) необходимы аэраторы, кислородные баллоны. В этом случае плотность посадки должна быть не более 10-12тыс. шт. на одну автоцистерну. Перевозку лучше проводить в ночные часы или с охлаждением воды до 13-16°С.
Соотношение воды при перевозке рыбы в зависимости от расстояния распределяется следующим образом: при внутрихозяйственных перевозках соотношение рыбопосадочного материала к воде принимают 1:3, а соотношение товарной рыбы - 1:2, при дальних перевозках плотность сокращается до соотношения 1:4, товарной- 1:3.
Зарыбление прудов мальками осетровых:
Доставленную молодь выдерживают в транспортной емкости, подавая в нее прудовую воду до уравнивания температуры воды в емкости и прудовой воды.
Пересчитывать доставленную молодь тотчас после транспортировки не рекомендуется во избежание ее дополнительного отхода. Для адаптирования молоди к местной воде и искусственным кормам полезно временно поместить ее в бассейны с проточной водой и начать регулярное кормление. Определение точной численности привезенной молоди позволит фермеру-рыбоводу рассчитать объем задаваемых кормов, тем самым, предотвратив неоправданные финансовые издержки. Естественных кормов в пруду с большой плотностью содержания рыб обычно мало, но они имеют значение в первые дни содержания в них молоди. В связи с этим следует контролировать биомассу бентических организмов.
Кормление молоди осетровых:
Приучать молодь к кормлению необходимо на всей площади пруда, для чего корм задается несколько раз в день в разные точки пруда на кормовые плиты. При этом следует проверять поедаемость пищи. Необходимо учесть, что для молоди массой до 100-200 г в кормах должно быть белков - 40-50%, жиров - 5, углеводов-10-12%.
С ростом молоди размеры частиц или гранул кормов увеличивают, что повышает эффективность роста рыб. В середине лета молодь может потреблять уже кильку, тюльку, бычков, атерину.
Наиболее эффективный способ кормления предполагает наличие кормовых таблиц, когда суточная доза корма выдается в зависимости от общей и средней индивидуальной массы рыбы, температуры воды и других условий выращивания. Ориентировочный расход кормов по месяцам соответствует указанному
В условиях фермерских хозяйств рекомендуется задавать корм, состоящий из следующих компонентов, %: фарш из отходов рыбы (карпа и толстолобиков) свежий - 35, фарш из мороженой рыбы -35, комбикорм карповый - 20, гидролизные дрожжи - 5, фосфатиды - 4, премикс форелевый - 1. Кормовой коэффициент такой смеси равен 7.
При плохом потреблении кормов следует увеличить проточность воды в прудах, проверить качество корма, произвести известкование кормовых мест.
Зимовка:
Зимовка проводится в тех же прудах, что и выращивание, предварительно подсушенных в течение двух-трех дней, обработанных хлорной и обычной известью с последующей промывкой. Для предотвращения промерзания водопадающих каналов их укрывают на зиму камышовыми матами. Можно продолжать кормить осетровых зимой, снизив рацион в 3-5 раз. Плотность их посадки на зиму при пятисуточном водообмене - 20 т/га. Отход осетровых за зимовку в среднем составляет 10%. Содержание кислорода в зимний период должно быть не ниже 5-6 мг/л (Дорохов С.М. 1981).
2.3.5 Защита русского осетра
В целях сохранения численности осетровых и увеличения пропуска их производителей на нерестилища Федеральным агентством по рыболовству с 2001 года в Волго-Каспийском районе введен сезонный запрет на промысел всех видов рыб с 11 мая до 31 августа. На других банках дельты Волги промысел запрещен с 26 мая по 31 августа.
В реке Волге с 2002 года запрещен специализированный промысел осетровых и разрешен их прием для целей воспроизводства, научных исследований и промышленной переработки. Все виды осетровых используются исключительно для целей воспроизводства и научных исследований. На участке реки Волги от города Астрахани до Волгоградской плотины ГЭС промысел всех видов осетровых запрещен. Анализ существующих данных показывает, что в Каспийском море исходя из современного состояния общих запасов осетровых при рациональном ведении рыбного хозяйства еще имеется возможность сохранить и в дальнейшем увеличить численность популяций (Мухачев И.С. 1989).
Охранный статус. Русский осетр включен в Красную книгу МСОП.
ГЛАВА 3. БЛОК-СХЕМЫ ИМИТАЦИОННОЙ МОДЕЛИ ДИНАМИКИ ЧИСЛЕННОСТИ ПОПУЛЯЦИИ РУССКОГО ОСЕТРА
Динамика численности русского осетра зависит от воздействия различных факторов. На основе влияния этих факторов построена блок-схема (рис.2.). Главными факторами являются: антропогенные, биотические, абиотические.
А в частности:
Соленость воды.
Соленость воды является переменным фактором влияния, так как в различных местах обитания, и по ходу миграции, соленость воды меняется. В основном соленость влияет на рост и развитие мальков осетра. Также влияет на распространение, в зонах с высокой соленостью русский осетр не обитает.
Температура.
Температура воды также является переменным фактором, так как в течение года и в различных местах температура меняется. Температура влияет на рост и развитие, также на распространение.
Освещенность.
Стабильная повышенная освещенность влияет на рост, и нормальное развитие мальков осетра, что в свою очередь влияет на численность русского осетра. Освещенность также является переменным фактором.
Насыщенность кислородом.
У осетра отмечается четкая видовая специфичность как в отношении минимального количества кислорода, растворенного в воде. Так, с увеличением содержания кислорода в определенном диапазоне концентраций происходит ускорение эмбриогенеза. Дальнейшее увеличение содержания кислорода приводит к замедлению развития зародышей и углублению образующихся аномалий.
От концентрации кислорода в воде зависит их жизнедеятельность. При уменьшении его ниже определенных границ падает интенсивность питания и использования пищи на рост, в результате чего замедляется рост.
При недостаточном содержании кислорода в воде снижается устойчивость к неблагоприятным факторам внешней среды, в том числе к промышленным и бытовым загрязнениям. Длительное пребывание в воде с недостаточным содержанием кислорода понижает активность, резко снижает устойчивость к возбудителям болезней.
Гидрологический уровень.
Гидрологический уровень влияет на распространение русского осетра. В основном осетр водится в глубоководных зонах. Этот фактор также является переменным.
Межвидовая конкуренция.
Межвидовая конкуренция влияет на численность, и распространение, так как русский осетр в основном конкурирует за питание.
Отношения «хищник – жертва».
Эти отношения являются основным параметром, так как они создают основу питания русского осетра, и влияют на рост, развитие и распространение.
Паразитизм.
Влияет на рост и развитие, как мальков, так и взрослых особей.
Промысел.
Является основным лимитирующим фактором в настоящее время. Это происходит, потому что он не угнетает, а полностью изымает особей из жизненного цикла.
Загрязнение среды обитания.
Загрязнение среды обитания более всего влияет на рост и развитие мальков осетра. В результате загрязнений происходят изменения генетического фонда еще на личиночной стадии, и рыбы уже изначально рождаются с уродствами и патологиями. Также загрязнения влияют на другие факторы влияния, такие как температура, мутность, содержание солей, кислорода и др.
Гидростроительство.
Гидростроительство на реках привело к уменьшению численности вида, так как оно влияет на распространение, а точнее не дают рыбам подняться на нерест, для размножения.
ЗАКЛЮЧЕНИЕ
Численность русского осетра зависит от действия множества факторов.
Проведя исследования по воздействию этих факторов можно сказать, что русский осетр соленоустойчивый вид, так как уже в периоды, когда его вес составляет 18 – 20 грамм, он выдерживает соленость прядка 12‰. Также можно сказать, что он относится к теплолюбивым, основная зона его распределения в диапазоне выше 20°C. Это прослеживается с самого рождения мальков, при температурах выше 20°C, наблюдается стабильный рост и нормальное усвоение пищи.
По отношению к кислороду, при его недостатке наблюдается увядание, и замедление развития, но чрезмерное содержание кислорода в воде может привести к летальному исходу. При оптимальном содержании кислорода в воде прослеживается, стабильный рост, активная жизнидеятельнось, и нормальное питание. Высокое содержание углекислого газа в воде прямо пропорционально связанно с содержанием кислорода, и косвенно воздействует на увядание русского осетра.
Русский осетр в основном обитает в мелководных зонах, но при понижении уровня моря, он мигрирует в более глубоководные зоны.
Так же как и другие виды, русский осетр вступает в межвидовую конкуренцию, за питание. И также он подвержен воздействию различных паразитов.
Отрицательным качеством этого вида является, долгий цикл развития, половая зрелость у этого вида наступает не ранее 10 лет, что сказывается на его воспроизводстве, хотя количество икринок при икрометании довольно велико.
Основная проблема воспроизводства этого вида заключается в антропогенном воздействии. Гидростроительство на реках преграждает путь к основному нерестилищу, и много особей погибает, не дойдя до него.
И на настоящий день самой важной проблемой выживания русского осетра является его вылов, так как он считается очень ценным промысловым объектом, как и другие виды осетровых.
Из этого всего следует, что хотя русский осетр, является устойчивым к различным факторам, но антропогенная проблема на данный момент считается самой важной для дальнейшего развития численности популяции.
1. Изучив общую характеристику русского осетра можно сказать, что у него довольно долгий цикл развития. При достижении половой зрелости его вес может достигать более 140 кг., а размер более 2х м. в длину. Существуют яровые и озимые формы осетра. Основной нерестовой рекой является Волга, а обитает в Черном, Азовском и Каспийском море.
2. По воздействию различных факторов: русский осетр является теплолюбивым, солеустойчивым, также легко переносит другие факторы. Лимитирующими факторами являются - антропогенные.
3. Имитационная модель динамики численности русского осетра зависит от воздействия множества факторов, которые показывает блок-схема (рис.2).
СПИСОК ЛИТЕРАТУРЫ
1. Берг Л.. Яровой и озимой расы мигрирующих рыб. АН СССР Press. 1934. –С: - 711-732.
2. Богатова И.Б. Рыбоводная гидробиология М., 1980. – С: 34.
3. Бурмаков Г.Т., Моисеев Н.Н. Прудовое рыбоводство. Кемерово, 1981. – С: - 81.
4. Вишнякова М.А., Брудастова Р.И. Гидротехнические сооружения рыбоводных хозяйств. М., 1985. – С: - 56.
5. Вишнякова М.А., Брудастова Р.И. Биология пресноводных рыб и методы их вылова. М., 1989. – С: - 77.
6. Гербильский. Васильева Л.М. Биологические и технологические особенности аквакультуры осетровых в условиях нижнего Поволжья. - Астрахань, 2000. - 190 с.
7. Данилов Ю.М. Аэрация воды рыбохозяйственных водоемов. М., 1980.
8. Детлаф, Таас Гинзбург, О. И. Shmalgauzen. Осетрина развития. Наука. Москва. 1981.- 224 с.
9. Денисов А.И. Рыбоводство на водохранилищах. М., 1978.- 129с.
10. Дорохов С.М. Прудовое рыбоводство. М., 1981.- 38с.
11. Иванов А.П. Рыбоводство в естественных водоемах. М., 1988. -45с.
12. . Исаев А.И. Разведение рыбы в оросительных каналах и на рисовых полях.М., 1988. – 80с.
13. Карпевич Л.Ф. Теория и практика акклиматизации водных организмов. М., 1975.-93с.
14. Козлов В.И., Абрамович Л.С. Товарное осетроводство. М., 1986.-23с.
15. Козлов В.И. Ирригация и рыба. Ставрополь, 1977.-67с.
16. Королева В.А. Состояние и перспективы развития аквакультуры. М., 1985.- 46с.
17. Мельников В.Н. Биотехнические основы промышленного рыболовства. М., 1983.- 86с.
18. Милн П. Морские хозяйства в прибрежных водах. М., 1978.- 96с.
19. Михеев В.П. Садковое выращивание товарной рыбы. М., 1982.-104с.
20. Мухачев И.С. Озерное рыбоводство. М., 1989.- 98с.
21. Никоноров А.М., Хоружая Т.А., Бражникова Л.В., Жулидов А.В. Мониторинг качества вод: оценка токсичности. С.-Петербург. Гидрометеоиздат. 2000. 150 с
22. Никольский Г.В. Экология рыб. М., 1974. – 47с.
23. Привезенцев Ю.А. Использование теплых вод для разведения рыбы. М.,1986.999.- С. 25-32.
24. Ходоревская, RP, Е. В. Красиков, Ф. Довгопол и О. Журавлева, Ихтиологический мониторинг состояния осетра в Каспийском море. В: мониторинге биоразнообразия. Москва. 1997. 159-164.
ПРИЛОЖЕНИЕ
Корреляционно-регрессионный анализ
Построим график, отложив вдоль оси абсцисс содержание х, а вдоль оси ординат- у. Тогда каждой паре значений х и у на графике будет соответствовать определенная точка. По характеру расположения точек можно предположить существование нелинейной функции между х и у.
| х | 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 15 | 20 |
| у | 14 | 30 | 55 | 112 | 200 | 361 | 594 | 830 | 1074 | 1268 | 1360 | 1428 | 1450 | 1490 | 1499 |
Рис.1 График экспериментальных данных
Из данного графика мы можем определить, что приведенное уравнение является логистическим уравнением Ферхюльста(1838), описывающим рост популяции в условиях внутривидовой конкуренции.
Если δ- коэффициент пропорциональности;
У - численность популяции в момент х;
r - коэффициент роста;
К - предельная плотность популяции
начальное условие: при х=0 численность особей у=![]()
отсюда логистическое уравнение:
![]()
![]()
![]()
![]()
![]()
При х=1; у=30; ![]() =14;К=1499 (из данного графика):
=14;К=1499 (из данного графика):
![]()
![]()
![]()
![]()
| х | 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 15 |
| ŷ | 14.764 | 30.338 | 61.657 | 122.608 | 233.99 | 415.97 | 665.02 | 934.42 | 1160.87 | 1314.24 | 1403.53 | 1451 | 1475.025 | 1495.454 |
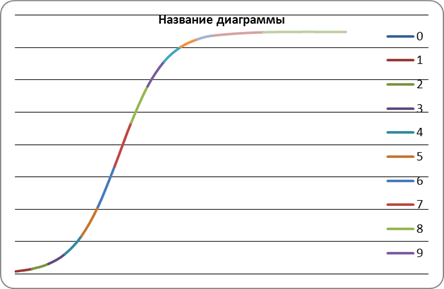
Рис.2 График теоретической зависимости
Остаточная дисперсия:
![]()
![]()