| Похожие рефераты | Скачать .docx | Скачать .pdf |
Реферат: Экология популяции
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ
ФЕДЕРАЛЬНОЕ АГЕНТСТВО ПО ОБРАЗОВАНИЮ
ГОУ ВПО «СЫКТЫВКАРСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ»
ХИМИКО-БИОЛОГИЧЕСКИЙ ФАКУЛЬТЕТ
КАФЕДРА ЭКОЛОГИИ
РЕФЕРАТ
Экология популяции
Проверила: Колесникова А.А.
Выполнила: Осташова Е.В.
Сыктывкар
2008
Содержание
Понятие популяции в экологии
Структура популяции и её виды
Пространственные подразделения популяции
Численность и плотность популяций
Место популяции в иерархии биологических систем
Список использованной литературы
Понятие популяции в экологии
Популяцией в экологии называют группу особей одного вида, находящихся во взаимодействии между собой и совместно населяющих общую территорию.
Слово «популяция» происходит от латинского «популюс» - народ, население. Экологическую популяцию, таким образом, можно определить как население одного вида на определённой территории.
Члены одной популяции оказывают друг на друга не меньшее воздействие, чем физические факторы среды или другие обитающие совместно виды организмов.
В популяциях в той или иной степени проявляются все формы связей, характерные для межвидовых отношений, но наиболее ярко выражены мутуалистические (взаимно полезные) и конкурентные.
Специфические внутривидовые взаимосвязи – это отношения, связанные с воспроизводством, между особями разных полов и между родительским и дочерним поколениями.
При половом размножении обмен генами превращает популяцию в относительно целостную генетическую систему. Если перекрёстное оплодотворение отсутствует и преобладают вегетативное партеногенетическое или другие способы размножения, то генетические связи слабее и популяция представляет собой систему клонов, или чистых линий, совместно использующих среду. Такие популяции объединены в основном экологическими связями. Во всех случаях в популяциях действуют законы, позволяющие использовать ограниченные ресурсы среды таким образом, чтобы обеспечить оставление потомства. Достигается это в основном через количественное изменение населения. Популяции многих видов обладают связями, позволяющими им регулировать свою численность.
Поддержание оптимальной в данных условиях численности называют гомеостазом популяции. Гомеостатические возможности популяций по-разному выражены у различных видов. Осуществляются они также через взаимоотношения особей.
Таким образом, популяции как групповые объединения обладают рядом специфических свойств, которые не присущи каждой отдельно взятой особи. Групповые особенности – это основные характеристики популяций. К ним относятся: 1) численность – общее количество особей на выделяемой территории; 2) плотность популяции – среднее число особей на единицу площади или объёма занимаемого популяцией пространства; 3) рождаемость – число новых особей, появившихся за единицу времени в результате размножения; 4) смертность – показатель, отражающий количество погибших в популяции особей за определённый промежуток времени; 5) прирост популяции – разница между рождаемостью и смертностью; прирост может быть как положительным, так и отрицательным; 6) темп роста – средний прирост за единицу времени.
Популяции свойственна определённая организация. Распределение особей по территории, соотношения групп по полу, возрасту, морфологическим, физиологическим, поведенческим и генетическим особенностям отражают структуру популяции. Она формируется, с одной стороны, на основе общих биологических свойств вида, а с другой – под влиянием абиотических факторов среды и популяций других видов. Структура популяций имеет, следовательно, приспособительный характер. Разные популяции одного вида обладают как сходными особенностями структуры, так и отличительными, характеризующими специфику экологических условий в местах их обитания.
Таким образом, кроме адаптивных возможностей отдельных особей население вида на определённой территории характеризуется ещё и приспособительными чертами групповой организации, которые являются свойствами популяции как надиндивидуальной системы. Адаптивные возможности вида в целом как системы популяций значительно шире приспособительных особенностей каждой конкретной особи.
Зародилось и успешно функционирует целое научное направление – демэкология (от греч. demos – народ), рассматриваемое как раздел общей экологии, изучающий структурные и функциональные характеристики, динамику численности популяции, внутрипопуляционные группировки и их взаимоотношения, выясняющий условия, при которых формируются популяции, и др. При этом приоритетной для демэкологии проблемой являются биотические взаимоотношения.
Структура популяции и ее виды
Любой популяции присуща определенная организация. Распределение особей по территории, соотношение групп особей по полу, возрасту, морфологическим, физиологическим, поведенческим и генетическим особенностям отражают соответствующую структуру популяции: пространственную, половую, возрастную и т.д. Структура формируется, с одной стороны, на основе общих биологических свойств видов, а с другой — под влиянием абиотических факторов среды и популяций других видов. Поэтому важно подчеркнуть откровенно приспособительный характер структуры популяции.
Возрастная и половая структуры популяции. Возрастная структура популяции, т.е. соотношение в ней разных возрастных групп, определяется особенностями жизненного цикла вида и внешними условиями.
В любой популяции можно условно выделить три экологические группы: предрепродуктивную, репродуктивную, пострепродуктивную. К предрепродуктивной относится группа особей, возраст которых не достиг способности к воспроизведению; репродуктивная — группа, способная воспроизводить новые особи; наконец, пострепродуктивная — особи, которые по ряду причин утратили способность участвовать в воспроизведении новых поколений. По отношению к общей продолжительности жизни длительность этих возрастов сильно меняется у разных видов. Для человека эти три возраста приблизительно одинаковы.
Имеются виды с очень простой возрастной структурой популяций, которые состоят практически из представителей одного возраста. Так, все однолетние растения весной находятся в проростках, затем почти одновременно зацветают, дают семена и к осени отмирают. Уязвимость таких популяций крайне высока: если в период развития наступают, например, заморозки, происходит массовая гибель особей. Напротив, в благоприятной ситуации такая популяция может дать взрыв численности (саранча, грызуны).
В популяции со сложной возрастной структурой представлены все возрастные группы, одновременно живут несколько поколений. Так, в стадах слонов, например, есть и новорожденные, и подростки, и молодые крепнущие животные, и способные к размножению самцы и самки, и старые особи. Такие популяции не подвержены резким колебаниям численности. Экстремальные внешние условия могут изменить их возрастной состав за счет гибели наиболее слабых, но самые устойчивые возрастные группы выживают и затем восстанавливают структуру популяции.
Очевидно, что человек, рассматриваемый как биологический вид, имеет сложную структуру популяций.
Половые группировки внутри популяций формируются на базе различной морфологии (формы и строения тела) и экологии различных полов. Отличие самцов от самок затрагивает не только строение и функцию половой системы, но и морфологию в целом (рога у самцов и отсутствие их у самок; крылатые самцы и бескрылые самки у некоторых насекомых; яркое оперение самцов и скромное у самок, и т.д.). Часто встречается различие самцов и самок по характеру и виду пищи. Так, у многих комариных самцы питаются нектаром или соком растений, а самки — кровью жертв. Разный пищевой рацион самцов и самок имеет место у ряда млекопитающих, птиц, рыб.
Соотношение возрастных и половых групп, которые можно отнести к статическим показателям популяции, определяет многое в общей жизнеспособности и темпах роста популяции, является важной характеристикой ее структуры. Тщательный и постоянный анализ возрастного и полового состава популяций — главное условие для прогнозирования численности тех видов, которые человечество использует в дикой природе или с которыми вынуждено бороться.
Пространственная структура популяций. Пространство, которое занимает популяция, дает ей необходимые для жизни условия. Однако каждая конкретная территория способна прокормить лишь определенное число особей. При этом очевидно, что на степень использования доступных природных ресурсов влияет не только общая численность популяции, но и размещение особей в пространстве.
Изредка в природе встречается почти равномерное упорядоченное распределение особей на занимаемой территории, например в чистых зарослях некоторых растений. Однако в силу неоднородности занимаемого пространства, а также некоторых особенностей биологии видов чаще всего члены популяции распределяются в пространстве неравномерно. При этом существуют два крайних варианта неравномерного размещения членов популяции: 1) четко выраженная мозаичность с незанятым пространством между отдельными скоплениями особей (например, гнездовья грачей в рощах или парках); 2) распределение случайного, диффузного типа, когда члены популяции более или менее независимы друг от друга и обитают в относительно однородной для них среде (например, размещение мучных хрущаков в муке).
Отметим, что между указанными вариантами неравномерного размещения существует множество переходов. В то же время тип распределения в занимаемом пространстве в каждом конкретном случае оказывается приспособительным, поскольку позволяет оптимально использовать имеющиеся ресурсы.
Растения чаще всего распределены крайне неравномерно, образуя при этом более или менее изолированные группы, скопления, которые называются субпопуляциями. Они имеют отличия в числе особей, плотности, возрастной структуре и протяженности. Напротив, у животных, благодаря их подвижности, способы упорядочивания территориальных отношений более разнообразны по сравнению с растениями. При этом внутрипопуляционное распределение у высших животных регулируется системой инстинктов. Такие инстинкты, которые способствуют поддержанию размещения по территории отдельных особей или групп в популяциях, характерны для птиц, млекопитающих, пресмыкающих, ряда рыб.
По типу использования пространства все подвижные животные подразделяются на оседлых и кочевых.
Оседлые животные отличаются инстинктами привязанности к своему участку, стремлением вернуться на хорошо знакомую территорию (если произошло вынужденное переселение). Такое «чувство дома» получило название «хоминг» (от англ. home — дом). Яркий пример хоминга — занимание одной и той же парой скворцов «своего» скворечника в течение ряда лет.
Оседлому образу жизни присущи существенные биологические преимущества. В частности, на знакомой территории облегчается свободная ориентация, животное тратит меньше времени на поиски корма, быстрее находит укрытие от врага, а также может при необходимости создавать запасы пищи (белка, сурок, полевая мышь). В то же время оседлый образ жизни угрожает быстрым истощением пищевых ресурсов, если, например, плотность популяции становится чрезмерно высокой.
Даже в случае территориального обособления члены популяции поддерживают друг с другом определенную связь. Это обеспечивается при помощи системы различных сигналов и прямых контактов на границах владений.
Различают два типа активности территориального поведения животных: первый направлен на обеспечение собственного существования (поиск пищи, устройство убежищ и т.п.), второй — на установление отношений с соседями (мечение и охрана своих участков). При этом применяются разные способы для закрепления участка: от прямой агрессии по отношению к чужаку, до ритуального поведения, демонстрирующего угрозу, а также специальных сигналов и меток, которые свидетельствуют о занятости территории. Следует подчеркнуть, что прямая агрессия с нанесением конкуренту повреждений встречается весьма редко.
На нейтральной территории агрессивный инстинкт угасает. Частичное перекрывание индивидуальных «владений» служит способом поддержания контактов между членами популяции. При этом соседи, как правило, поддерживают устойчивую обоюдовыгодную систему связей, например, совместную защиту от врагов, взаимное предупреждение об опасности (сороки в лесу).
Территориальное поведение животных особенно ярко выражено в период размножения. По его окончании у многих видов распределение по индивидуальным участкам сменяется групповым образом жизни с иным типом поведения. Так, после вылета птенцов большинство воробьиных птиц объединяются в стаи, которые совершают кочевки.
Для оседлых видов животных все варианты общей пространственной структуры популяций обычно сводятся к четырем основным типам: диффузному, мозаичному (рассмотрены ранее), пульсирующему и циклическому.
Для популяций, характеризующихся резкими колебаниями численности, характерен пульсирующий тип пространственной структуры. Известно, что в период резкого падения численности некоторые животные собираются на наиболее благоприятных для жизни участках. Так, например, полевки-экономки в лесостепи в засушливые годы заселяют в первую очередь заболоченные берега озер.
Циклический тип пространственной структуры популяций оседлых животных характеризуется закономерным попеременным использованием территории в течение года, например летом и зимой. При таком типе использования пространства сохраняется баланс между потреблением кормов и их ежегодным возобновлением.
Кочевой образ жизни имеет перед оседлым определенные преимущества. Прежде всего, кочевые животные не зависят от запасов корма на конкретной территории. Однако постоянные передвижения одиночных особей способствуют учащению гибели от хищников. Именно поэтому кочевой образ жизни, как правило, свойствен стадам и стаям. При этом территории перемещения многих видов могут быть весьма большими. Так, табуны зебр в Серенгети в период сухого сезонакочуют на участке в 400—600 км2 . На данной территории имеются определенные места отдыха, пастьбы и водопоев, используемые регулярно в определенные сроки.
Многообразие конкретных форм проявления территориальных взаимоотношений животных ученые располагают в постепенно усложняющийся ряд. При этом система неперекрывающихся охраняемых участков оседлых особей — это самая простая пространственная структура популяций; более сложная — система перекрывающихся территорий, при которой усиливаются внутривидовые контакты. Наконец, групповое использование одних и тех же участков, возможное только на основе строго упорядоченных отношений внутри группы.
Этологическая структура популяции. Систему взаимоотношений между членами одной популяции называют этологической, или поведенческой структурой популяции.
Формы совместного существования особей в популяции весьма различны. Ниже рассмотрены наиболее характерные.
Одиночный образ жизни характерен для многих видов (например, ежи, сомы, щуки и т.д.), но лишь на определенных стадиях жизненного цикла. Поэтому абсолютно одиночного существования организмов в природе не встречается, иначе погибли бы соответствующие популяции.
У видов с преимущественно одиночным образом жизни часто возникают временные скопления — в местах зимовок, а также в период перед размножением. Так, божьи коровки осенью образуют целые гроздья в сухой подстилке возле пней и комлей деревьев.
Проживание в группе себе подобных отражается на протекании многих физиологических процессов в организме животного. У искусственно изолированных особей заметно меняется уровень метаболизма (обмена веществ), быстрее тратятся резервные вещества, не проявляется целый ряд инстинктов и ухудшается общая жизнеспособность. Под эффектом группы понимают оптимизацию физиологических процессов, ведущую к повышению жизнеспособности особей при их совместном существовании.
Эффект группы проявляется как реакция отдельной особи на присутствие других особей своего вида. Так, у овец вне стада резко учащаются пульс и дыхание, а при виде приближающегося стада эти процессы нормализуются, и овца успокаивается. Известно также, что для выживания африканских слонов стадо должно состоять по крайней мере из 25 особей. Эффект группы состоит также в ускорении темпов роста животных, повышении плодовитости, более быстром образовании условных рефлексов, повышении средней продолжительности жизни индивидуума. В группах животные часто способны поддерживать оптимальную температуру (при скучивании, в гнездах, ульях). У многих животных вне группы не реализуется плодовитость. Так, голуби некоторых пород не откладывают яйца, если не видят других птиц. Достаточно поставить перед самкой зеркало, чтобы она приступила к яйцекладке.
Эффект группы не проявляется у видов, ведущих одиночный образ жизни. Если таких животных искусственно заставить жить вместе, у них повышается раздражительность, учащаются столкновения, повышаются энергозатраты на поддержание жизнедеятельности. Так, ушастые ежи в группе повышают потребление кислорода до 134% по сравнению с содержащимися в одиночестве.
Согласно В.А. Радкевичу, в явлениях эффекта группы и особенно фазовой изменчивости огромную роль играют гормональные механизмы. Так, особые эндокринные железы насекомых — прилежащие тела — вырабатывают гормон, который стимулирует и управляет переходом особи из одиночной фазы в стадную.
Биологи считают, что самое сильное проявление эффекта группы свойственно общественным насекомым (пчелам, муравьям, термитам). Не обладая способностью длительно существовать в одиночку (что заложено у них в генетической программе), эти насекомые выработали сложную систему сигнализации, которая способствует сохранению их особей во времени и пространстве.
Изложенное позволяет понять, почему предъявляются повышенные требования при формировании групп космонавтов, отрядов специального назначения, экипажей, которые должны длительно находиться в замкнутом пространстве, либо длительно общаться друг с другом. При удачном подборе в таком коллективе явно проявляется «эффект группы» и он успешно справляется с поставленной задачей.
Семейный образ жизни резко усиливает связи между родителями и их потомством. Известное проявление этого — забота одного из родителей об отложенных яйцах или кормление самцом самки. При этом заботы о птенцах продолжаются до поднятия их на крыло, а у ряда крупных млекопитающих (медведей, тигров) детеныши воспитываются в семейных группах в течение нескольких лет, до наступления их половой зрелости. В зависимости от того, кто из родителей берет на себя уход за потомством, различают семьи отцовского, материнского и смешанного типа. Отметим, что в семьях с устойчивым образованием пар обычно оба родителя принимают участие в охране и выкармливании молодняка.
При семейном образе жизни территориальное поведение животных выражено наиболее ярко. Присущие ему различные сигналы, маркировка, ритуальные формы угрозы и даже прямая угроза (часто со стороны и самца и самки) обеспечивают владение участком, размеры и пищевая емкость которого достаточна для выкармливания потомства.
Колония. Будучи групповым поселением оседлых животных, она может существовать как длительно, так и возникать лишь на период размножения (грачи, чайки, гагары и т.п.).
Значительно более сложная форма колонии — поселения животных, в которых отдельные их жизненные функции выполняются сообща; Это в свою очередь повышает вероятность выживания отдельных особей. Так, тревога, поднятая любой заметившей хищника птицей, мобилизует остальных и им сообща удается его отогнать.
Некоторые общественные насекомые — пчелы, муравьи, термиты организуют весьма сложные колонии — семьи. Здесь насекомые выполняют сообща много основных функций: защиты, размножения, обеспечения кормом себя и потомства, строительства и т.п., для чего осуществляют обязательное разделение труда и специализацию отдельных особей, в том числе разных возрастных групп. При этом члены колонии постоянно обмениваются информацией друг с другом.
Характерно, что по мере усложнения колониального объединения поведение, а иногда даже физиология и строение отдельной особи все в большей степени подчиняются интересам всей колонии.
Стая. Это временное объединение животных одного вида (насекомых, птиц, рыб, реже млекопитающих и др.), связанное с общностью места обитания или размножения. Стайность облегчает выполнение каких-либо функций в жизни вида, например, защиты от врагов, добычи пищи, миграции.
Исходя из способа координации действий, стаи подразделяются на два вида: 1) без выраженного лидера (обычно у рыб); 2) с лидерами, на которых ориентируются остальные особи (стаи крупных птиц и млекопитающих, например волков). Волчьи стаи образуются зимой для групповой охоты. В этом случае зверям удается справиться с крупными копытными (например взрослым лосем), охота на которых в одиночку часто заканчивается гибелью самого хищника. В процессе групповой охоты вожак стаи «организует» засады, захват жертвы в кольцо и другие действия, что требует согласованности и координации действий всех членов стаи.
Стадо — это группа диких или домашних животных одного вида, обитающая на какой-либо территории (например, стадо оленей) или акватории. В стаде осуществляются все основные функции жизни: добывание корма, защита от хищников, миграции, размножение, воспитание молодняка и т.п. При этом основу группового поведения животных в.стадах составляют взаимоотношения доминирования(главенства) — подчинения, которые обусловлены индивидуальными различиями между особями.
Иерархически организованному стаду свойствен закономерный порядок перемещения, определенная организация при защите, расположении на местах отдыха и т.п. Так, при передвижении'стада павианов в центре, в наибольшей безопасности, находятся самки с детенышами или беременные, по краям вожаки, молодые самцы и неразмножающиеся самки. Впереди и позади стада располагаются крупные самцы, готовые отразить нападение хищника.
В биологическом аспекте смысл иерархической системы «доминирования-подчинения» заключается в создании согласованного поведения группы, которое выгодно для всех ее членов. Осуществив своеобразную «расстановку сил», животные уже не тратят лишней энергии на конфликты между отдельными особями, а группа в целом получает преимущества, подчиняясь наиболее сильным и опытным индивидуумам. Конечно, в экстремальных условиях (например, при отсутствии кормов), гибнут в первую очередь более слабые, подчиненные особи, но тем не менее под защитой группы они все же имеют больше шансов выжить, нежели в одиночку.
Внутривидовая конкуренция. В этом случае между особями сохраняется солидарность, так что они в состоянии размножаться и обеспечивать таким образом передачу свойственных популяции наследственных свойств. Очевидно, что внутривидовая конкуренция в корне отличается от межвидовой.
Внутривидвая конкуренция проявляется, например, в территориальном поведении, когда животное защищает место своего гнездования и известную площадь в его округе. Так, в период размножения самец колюшки охраняет определенную территорию, на которую, кроме своей самки, не допускает ни одной особи своего вида.
Другим проявлением внутривидовой конкуренции выступает существование социальной иерархии, которая характеризуется наличием особей доминирующих и подчиненных. У растений конкуренция касается главным образом света и воды. В первом случае растения при слишком большой густоте затеняют друг друга, что приводит к гибели их значительного числа. У деревьев формируется различный вид в зависимости от того, растут ли они в лесу или отдельно от других деревьев.
В ряде случаев внутривидовая конкуренция может приводить к дифференциации вида, когда последний распадается на несколько популяций, занимающих разные территории. Так, у саванной овсянки один экологический подвид размещается на прибрежных солончаках, другой — на сухих холмах.
У представителей одного и того же вида животных встречается и такое редкое явление, как каннибализм, т.е. поедание себе подобных. Он наиболее развит у хищных рыб-щук, окуней, трески, наваги и др. В условиях обостренной конкуренции за пищу или воду каннибализм проявляется подчас и у нехищных животных. Например, личинки майского жука, которые помещены в сухую почву, могут поедать друг друга.
Паразитирование на себе подобных, т.е. внутривидовой паразитизм, встречается нечасто, оно характеризует в основном отношения разных полов. Так, самки глубоководных рыб-удильщиков носят на себе значительно более мелких самцов, которые прирастают ртом к их телу и питаются как паразиты. Внутривидовой паразитизм имеет приспособительное значение, поскольку наличие «карманных» самцов исключает необходимость затраты энергии на встречу полов, а также снижает остроту конкуренции из-за пищи в условиях общего недостатка кормов на больших глубинах.
Пространственные подразделения популяций
Пространство или ареал, занимаемое популяцией, может быть различным как для разных видов, так и в пределах одного вида. Величина ареала популяции определяется в значительной мере подвижностью особей или радиусом индивидуальной активности. Если радиус индивидуальной активности невелик, величина популяционного ареала обычно также невелика (табл. 1).
Таблица 1
Величина радиуса индивидуальной активности животных и растений (по А.В. Яблокову, А.Г. Юсуфову, 1976)
| Вид | Радиус активности |
| Виноградная улитка (Helixpomaceae) | Несколько десятков метров |
| Сельдь (Clupea harengus) | Несколько сот километров |
| Песец (Alopex lagopus) | Несколько сот километров |
| Северный олень (Rangifer tarandus) | Более ста километров |
| Ондатра (Ondatra zibetica) | Несколько сот метров |
| Усатые киты (Mysticeti) | Несколько тысяч километров |
| Дуб(пыльца) (Quercus petraca) | Несколько сот метров |
У Растений радиус индивидуальной активности определяется расстоянием, на которое могут распространяться пыльца, семена или вегетативные части, способные дать начало новому растению. Во многих других случаях трофический ареал не совпадает с репродукционным. Так, несмотря на огромный трофический ареал белого аиста (Ciconia alba), обитающего в Европе, а зимой – в Африке, каждая пара птиц возвращается обычно в район своего старого гнезда, и популяции аистов, хотя и смешиваются на местах зимовок, но во время размножения занимают относительно небольшую территорию.
В зависимости от размеров занимаемой территории Н. П. Наумов (1963) выделяет три типа популяций: элементарные, экологические и географические (рис.1).
Элементарная, или микропопуляция, - это совокупность особей вида, занимающих какой-то небольшой участок однородной площади. В состав их обычно входят генетически однородные особи. Количество элементарных популяций, на которые распадается вид, зависит от разнородности условий среды обитания: чем они однообразнее, тем меньше элементарных популяций, и наоборот. Между элементарными популяциями всегда имеются некоторые отличия, проявляющиеся в генетическом своеобразии, фенологических особенностях, способности к накоплению питательных веществ, интенсивности обмена, в характере поведения, или каждая элементарная популяция морфофизиологически и этологически (поведенчески) специфична, различия между ними определяются их генетическим своеобразием и средой обитания. Однако нередко смешение особей элементарных популяций, происходящее в природе, стирает границы между ними.
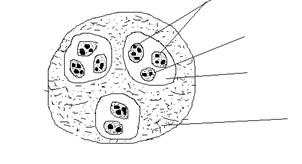
Рис. 1. Пространственное подразделение популяций (по Н.П. Наумову, 1963):
1 – ареал вида; 2 – 4 – соответственно географическая, экологическая, и элементарная популяции
Экологическая популяция формируется как совокупность элементарных популяций. В основном это внутривидовые группировки, слабо изолированные от других экологических популяций вида, поэтому обмен генетической информаций между ними происходит сравнительно часто, но реже, чем между элементарными популяциями. Экологическая популяция имеет свои особые черты, отличающие её в чём-то от другой соседней популяции. Так, белки (Sciurus vulgaris) заселяют различные типы леса, и могут быть чётко выделены «сосновые», «еловые», «пихтовые», «елово- пихтовые» и другие их экологические популяции.
Географическая популяция охватывает группу особей, населяющих территорию с географически однородными условиями существования. Географические популяции занимают сравнительно большую территорию, довольно основательно разграничены и относительно изолированы, Они различаются плодовитостью, размерами особей, рядом экологических, физиологических, поведенческих и других особенностей. Для географической популяции характерен генетический обмен, и хотя он может быть редким, но всё же возможен. При перекрёстном скрещивании особи каждой популяции приобретают общий морфологический тип, в чём-то несколько отличающийся от соседней географической популяции, с которой регулярного контакта нет. Например, узкочерепная полёвка занимает большой ареал. Она встречается в степных районах нашей страны и далеко на севере в зоне тундры. Географические популяции из тех и других районов имеют существенные отличия между собой по физиологии и размерам животных. Тундровые в отличие от степных более крупные, значительно раньше начинают размножаться, обладают более высокой плодовитостью и больше накапливают жира. Отличия настолько чётко выражены, что длительное время считали эти группы разными видами. Однако эксперименты показали, что обе формы полёвок легко скрещиваются и дают плодовитое потомство, следовательно, принадлежат к одному виду.
Границы и размеры популяций в природе определяются особенностями не только заселяемой территории, но в первую очередь свойствами самой популяции. Здесь всегда лежит степень её генетического и экологического единства. Раздробление вида на множество мелких территориальных группировок носит приспособительный характер к большому разнообразию местных условий, что увеличивает генетическое многообразие вида и обогащает его генофонд. Таким образом, наиболее общим правилом является то, что индивиды любого живого вида всегда представлены не изолированными отдельностями, а их определённым образом организованными совокупностями. Это правило было сформулировано в 1903 г. С.С. Четвериковым (1880 – 1959) и получило название правила объединения в популяции.
Численность и плотность популяций
Основными показателями структуры популяций является численность и распределение организмов в пространстве и соотношение разнокачественных особей. В связи с размерами ареала популяций может значительно изменяться и численность особей в популяциях.
Численность популяции – это общее количество особей на данной территории или в данном объёме. Зависит от соотношения интенсивности аразмножения (плодовитости) и смертности. В период размножения происходит рост популяции. Смертность же, наоборот, приводит к сокращению её численности.
Плотность популяции определяется количеством особей или биомассой на единицу площади либо объёма, Например: 400 деревьев на 1 га, 0,5 г циклопов в 1 м3 воды. Нередко важно различать среднюю плотность, т. е. численность или биомассу на единицу всего пространства, и удельную или экологическую плотность – численность или биомассу на единицу обитаемого пространства, доступной площади объёма, которые фактически могут быть заняты популяцией.
Плотность популяции отличается изменчивостью и зависит от её численности. При возрастании численности не наблюдается увеличение плотности лишь в том случае, когда возможно распределение популяции, расширение её ареала. Особи, составляющие популяции, имеют различные типы пространственного распределения, выражающие их реакции на различные влияния, например, добычу и благоприятные физические условия или конкурентные реакции. Различают три типа распределения или расселения особей внутри популяции: равномерное, случайное и групповое.
Равномерное распределение в природе чаще связано с острой конкуренцией между разными особями. Такой тип распределения отмечают у хищных рыб и у колюшек с их территориальным инстинктом и сугубо индивидуальным характером.
Случайное распределение имеет место только в однородной среде. Так на первых порах распределяется тля на поле. По мере её размножения распределение приобретает групповой или пятнистый (конгрегационный) характер.
Групповое распределение встречается наиболее часто. Так, в сосновом лесу деревья вначале расселяются группами, а в дальнейшем их размещение становится равномерным. Популяции групповое распределение обеспечивает более высокую устойчивость по отношению к неблагоприятным условиям по сравнению с отдельной особью. Животные, ведущие подвижный образ жизни, как правило, распределяются активно, что приводит к интенсивному перемешиванию популяций и стиранию границ между ними. Например, очень подвижные и активно перемещающиеся песцы, другие животные, птицы имеют огромные ареалы без резких границ между популяциями.
У пассивно передвигающихся и малоподвижных организмов, наоборот, популяции чётко разграничены даже на относительно небольшой территории. Таковы популяции наземных моллюсков, многих земноводных. Размеры ареала популяции зависят от величины особей, составляющей её. Мелкие особи занимают сравнительно небольшие ареалы, тогда как у видов с крупными особями они обширны. Вместе с тем это правило имеет много исключений. Так, территория, занимаемая популяцией прыткой ящерицы Lacerta agilis, может колебаться от 0,1 до нескольких гектаров.
Знание типа распределения организмов имеет большое значение при оценке плотности популяции методом выборки (в случае группового размещения площадь выборки должна быть большая). Возьмём n выборок. Среднее число особей в каждой выборке обозначим через m и получим рассеяние или дисперсию S2 по формуле:
S2
= ![]()
При равномерном распределении дисперсия S2 равна нулю, так как число особей в каждой выборке постоянно и равно среднему. В случае случайного распределения среднее m и дисперсия S2 равны. При групповом распределении рассеяние S2 выше среднего и разница между ними тем больше, чем сильнее тенденция животных к образованию скоплений.
Место популяции в иерархии биологических систем
Сложность представления о популяции определяется двойственностью её положения в рядах иерархически соподчинённых биологических систем, отражающих различные уровни организации живой материи (рис.2). С одной стороны, популяция включается в генетико-эволюционный ряд, отражающий филогенетические связи таксонов разного уровня как результат эволюции разных форм жизни:
организм → популяция → вид → род → … → царство
В этом структурно-эволюционном ряду популяция выступает как форма существования вида и её основная функция заключается в обеспечении устойчивого выживания и воспроизведения вида в данных конкретных условиях. В этом аспекте наиболее значимые свойства популяции характеризуются её генетической спецификой: общность и специфичность генофонда определяют адаптивность всех особей и популяции в целом к конкретным условиям среды. В то же время определённый уровень генетической гетерогенности особей в популяции предоставляет материал для микроэволюционного процесса в условиях меняющейся среды или возникшей по тем или иным причинам изоляции. В этом случае популяция выступает не только как форма существования вида, но и как элементарная единица его эволюционного преобразования. Этому аспекту популяционной биологии посвящено много исследований, составляющих одно из важнейших направлений современной теоретической биологии.
Одновременно с этим, представляя вид в конкретных условиях среды, популяция вступает в трофические и иные отношения с популяциями других видов, т. е. включается в состав конкретного биогеоценоза. Этот подход к популяционным системам отражает функционально-энергетический ряд различных уровней организации жизни:
организм → популяция → биогеоценоз → биосфера
В этом ряду популяция выступает в качестве функциональной субсистемы конкретного биогеоценоза; её функция – участие в трофических цепях – определяется видоспецифическим типом обмена. Устойчивое выполнение этой функции основывается на адаптивности популяции, способности к поддержанию популяционного гомеостаза, вследствие чего обеспечивается стабильное участие вида в биогенном круговороте. Этому аспекту существования популяционных систем в наибольшей степени подходит «функциональный» принцип определения масштаба популяции.
«Двойственная» природа популяционных систем заключается ещё и в биологической противоречивости их функций. Будучи, по определению, группировками особей одного вида в общих местах обитания, популяции состоят из организмов, максимально сходных по их биологическим свойствам и требованиям к среде. Это потенциально содержит в себе высокую вероятность возникновения острых конкурентных отношений между особями в популяции. Адаптации к снижению уровня конкуренции действуют в направлении дифференциации, разобщения особей в составе популяции. Но как целостная система популяция может эффективно функционировать только в условиях устойчивых контактов и взаимодействий особей, что связано с задачей их сближения, интегрированности в составе популяции. Разрешение этих противоречий – основа структурированности популяционных систем, поскольку поддержание оптимального соотношения между внутрипопуляционными процессами дифференциации и интеграции идёт на основе пространственной и функциональной неоднородности распределения и взаимосвязей особей в составе целостной популяции.

Рис.2. Положение популяции в структуре биологических систем биосферы (по И. А. Шилову, 1988).
Список используемой литературы
1. Ю.Л. Хотунцев. «Экология и экологическая безопасность»: Учеб. пособие для студ. высш. пед. учеб. заведений. – М.: Издательский центр «Академия», 2002, - 480 с.
2. Экология: Учебное пособие / Под ред. проф. В.В. Денисова. Серия «Учебный курс». – Ростов н /Д: Издательский центр «МарТ», 2002. – 640 с.
3. Степановских А.С. Экология: Учебник для вузов. – М.: ЮНИТИ-ДАНА, 2001. – 703 с.
4. Коробкин В.И., Передельский Л.В. Экология. – Ростов н /Д: изд-во «Феникс», 2001 – 576 с.
5. Шилов И.А. Экология: Учеб. для биол. и мед. спец. вузов. – М.: Высш. шк., 1997. – 512 с.
6. В.А. Радкевич. Экология. Краткий курс. Мн., «Вышэйш. школа», 1997. – 304 с.
Похожие рефераты:
Популяция северных морских котиков (Callorhinus ursinus) на острове Тюлений
Проблема перенаселённости Земли. История вопроса от Мальтуса до наших дней
Популяция и ее признаки. Классификация природных ресурсов
Взаимодействие животных как фактор эволюции
Особенности оценки продуктивности сельскохозяйственных животных
Активность основных карбокмипептидаз в тканях пренатально алкоголизированных крыс
Анализ продуктивности молодняка крупного рогатого скота симментальской и черно-пестрой пород