| Скачать .docx |
Реферат: Классификация карбонатных пород и биоседиментология как основа регионального фациального анализа
Классификация карбонатных пород и биоседиментология как основа регионального фациального анализа
Г.Д. Исаев
Биоседиментология - это наука, изучающая роль организмов в осадочном процессе.
Предметом биоседиментологии является индивидуальный организм в соотношении с окружающей био- и абиотической средой, а также процесс его жизнедеятельности во всем многообразии его проявлений от начальной фазы в осадочной среде до литификации продуктов его деятельности.
По составу скелет породообразующих организмов может быть самый разнообразный [1]: кальцитовый, арагонитовый, кремнистый, железистый, фосфатнокремнистый, целестиновый и др.
Доминируют среди породообразователей группы организмов, которые образуют скелет за счет интенсивной секреционной деятельности.
Биокомпоненты - это все, что остается в осадочном бассейне после жизни организма. Это и мягкие ткани, и твердые скелетные образования, и продукты жизнедеятельности, и следы их воздействия. Наименование биокомпонентов (как и биофаций) опубликованы автором ранее [2], как и в целом авторская терминология по биоседиментологии.
Среди биокомпонентов можно установить две большие группы: автохтонная и аллохтонная [2]. Степень устойчивости скелетов к разрушению является главной характеристикой при классификации биокомпонентов. Именно по такому признаку R.N. Ginsburq [3] установил шесть типов скелетных образований: массивный, ветвистый, камерный, сегментный, корковый, спикуловый.
В составе основных биокомпонентов выделяется пять групп [2]: скелеты, фрагменты скелетов, эрокла- сты, глобоиды и биокласты. Среди фрагментов устанавливается два вида: аллофрагменты и автофрагменты. Среди биокластов выделяется два вида: осколки без признаков переноса и обломки с элементами транспортировки. Глобоиды могут быть и аллохтонными, т.е. перемещенными внутри места обитания организмов и за его пределы, и автохтонными, т. е. находящимися на месте обитания организма.
Организмы активно воздействуют на грунт: уничтожают первичную слоистость осадка, нарушают структуру зерен, измельчают и растворяют материал, образуют трубки и норы, вносят фекалии, способствуют началу цементации, обесцвечивают осадки, уменьшают либо увеличивают содержание органического вещества, влияют на ход диагенеза. Особое значение имеет увеличение ОВ в осадках как источника в последующем первичного углеродистого материала.
По роли в образовании пород породообразующие организмы разделяются на две большие группы: созидающие, или архитекторы, и разрушающие. Среди первых тоже выделяются две большие группы: стабилизаторы и конструкторы. Стабилизаторами называются организмы, которые, поселяясь на мягкий или слаболи- тифицированный субстрат, интенсивной секреционной деятельностью ограничивают его подвижность. Конструкторы - это группа бентосных прикрепленных организмов, которые, поселяясь на скальном субстрате, своей секреционной деятельностью создают положительные формы подводного рельефа. Среди группы разрушителей выделяется два типа породообразователей: дестабилизаторы и деструкторы. Дестабилизаторы - это организмы, которые активной жизнедеятельностью разжижают субстрат, взмучивают илистый осадок, делити- фицируя его, переводя продукты биоэрозии в водную среду. Деструкторы - это организмы, которые поселяются не в илистом осадке, а в скальном, скелетном и в литифицированном субстрате, разрушая его до класти- ки, переводя продукты деструкции в водную среду.
Все организмы, относящиеся к типу «конструкторов», характеризуются только жестким прикреплением к грунту и отличаются от жесткоприкрепленных стабилизаторов типом скелетов: массивный, спикуловый (каркасный), корковый и камерный. Среди организмов- конструкторов устанавливается две группы: с наружным скелетом и с внутренним скелетом. В состав первой группы включают формы с различным расположением скелетопродуцирующей ткани: экзосоматические конструкторы имеют как массивный (кишечнополостные, водоросли, мшанки и др.), так и корковый (кишечнополостные, водоросли, мшанки, строматолиты) тип скелета; инсоматические конструкторы имеют только камерный тип скелета, например рудисты, фо- раминиферы, брахиоподы, усоногие раки, гастроподы (Vermetus) и др. Внутрискелетные формы конструкторов характеризуются доминированием форм с массивным и спикуловым типом скелета, причем среди последних только формы с признаками каркаса (например, губки, сфинктозоа).
Типизация органогенных построек основывается на различном характере сочленения скелетов в пространстве, доминирующей форме рифостроящих организмов, интенсивности абразии и биоэрозии, текстурных и структурных особенностях.
Среди дестабилизаторов выделено две группы организмов: 1) онфаунная группа организмов - это организмы, копающие, роющие, перерабатывающие илистый осадок; 2) инфаунная группа организмов - это ведущие активный образ жизни на поверхности осадка.
Мы наблюдали породы (биотурбилиты), практически полностью переработанные илоедами и камнеточцами.
В состав поверхностных или онфаунных дестабилизаторов отнесены все группы организмов, ведущих активный образ жизни в области жизнедеятельности надповерхностного уровня.
Деструкторы в геологическом прошлом менее всего изучены только потому, что большинство из них не имело скелета и не могло литифицироваться и сохраниться в ископаемом состоянии. В составе деструкторов устанавливаются две самостоятельные группы организмов по способу воздействия на субстрат: 1) сверлильщики; 2) кластаторы.
В основе системы первичных биогенных структур лежит классификация А.Ф. Эмбри и И.Е. Кловэна [5], детально изложенная и популяризированная в монографии Дж. Уилсона [6].
Авторская система биогенных структур [2], как было сказано выше, базируется на разработках зарубежных исследователей и предполагает деление на две большие группы: аллохтонные (биокластиты) и автохтонные (био-фитолиты).
Группа аллохтонных биогенных структур, или био- кластитов, состоит из шести видов структур: мадстоун, вакстоун, пакстоун, грейнстоун, рудстоун. Эта группа структур разделяется на три типа, каждый из которых делится на два вида. Мы не будем особенно углубляться в характеристику этого класса биогенных структур - она дана у Дж. Уилсона [6] и А.Ф. Эмбри и И.Е. Кловэна [5].
Тип автохтонных биогенных структур, или фитобиолитов, разделяется на три крупных класса: баунд- стоуны, или каркасные, инфлаутстоуны, или субкар- касные, и биотурбилиты. Два последних класса структур установлены автором на основе собственных наблюдений. Баундстоуны и инфлаутстоуны формируются организмами-архитекторами, могущими создавать жесткокаркасные структуры (организмы-стабилизаторы и конструкторы), а биотурбилиты - результат энергичного воздействия на скальный, илистый, слабо- литифицированный и скелетный субстрат организмов- дестабилизаторов и деструкторов.
В классе баундстоунов четыре биогенные структуры: бафлстоун, байндстоун, фреймстоун и лайтстоун. Критерий различия очень простой - тип скелета основного каркасообразующего биокомпонента [2]. Для бафлстоуна характерен ветвистый, для байндстоуна - корковый, для фреймстоуна - массивный и для лайт- стоуна - камерный тип скелета.
Лайтстоун (англ. lightstone) в переводе означает «легкая порода» [7]. Термин предложен автором для обозначения биогенных каркасных структур, образованных организмами камерного типа, т. е. имеющих раковины.
Гидродинамика лайтстоунов слабо исследована. Большое значение имеет плотность захоронения (насыщенные и плотные ориктоценозы), а также наличие или отсутствие спарита, ила, либо присутствие корковых, обрастающих скелетов, стабилизирующих илистый осадок в межкомпонентном пространстве. Чем выше энергетика, тем плотнее захоронение скелетов в лайтстоунах, тем больше спарита и скелетов.
Инфлаутстоун (англ. infloatstoune) - биогенная структура, состоящая из инситных скелетов, не сочленяющихся друг с другом в пространстве и не образующих жесткий органогенный каркас. Отличается от фла- утстоуна тем, что большинство скелетных образований находятся в положении роста и захоронены на месте жизнедеятельности организмов [7]. Инфлаутстоун подразделятся на два крупных класса структур: скелетные и фрагментарные [2].
Биотурбилиты - новый тип в группе автохтонных (фито-биолитов) карбонатных пород. Это породы, образованные организмами-деструкторами и организмами-дестабилизаторами [7]. Это общее название для всех пород, испытавших существенное (до разрушения) воздействие биотурбаторов, различного таксономического состава. Среди этих организмов (кластато- ров, по автору) выделяются две группы: 1) организмы, скелетная часть которых распадается на элементы наподобие скелетов сегментного типа (например, Halimeda и альционарии); 2) организмы с внутренним сегментным и спикуловым скелетом. В результате деятельности деструкторов образуется два класса биогенных структур биотурбилитов: эрогермные и эрокласт- ные. Четыре другие класса биотурбилитов формируются организмами-дестабилизаторами: 1) на границе осадок - водная среда; 2) внутри илистого грунта.
Деятельность биотурбаторов внутри илистого либо песчаного осадка обычно носит упорядоченный, системный характер, поэтому эту группу биотурбилитов мы называем группой системных биотурбилитов. В их составе три класса: вихревые, форонидные (или трубчатые) и сложно связанные (или аэдисные).
Биогенные текстуры еще менее изучены, чем биогенные структуры.
Если понимать текстуру как соотношение в пространстве участков породы, имеющих моноструктур- ное, однородное, монокомпонентное строение, то большинство органогенных построек типа биогерм, биостром будет иметь одну текстуру: массивную - однородную с равномерными характеристиками распределения компонентов в трех измерениях.
В ядерной части органогенных сооружений доминируют различные модификации массивной - однородной - и иловастных текстур. К группе биоаккуму- лятивных текстур кроме двух последних следует относить и еще две: биоритмитная и калиптровая. Биорит- митная макротекстура образуется в областях, где «давление» или влияние настоящих осадков на биоаккумуляцию достаточно сильно и выражается в последовательном чередовании биогенных и механокластических отложений, а также сменой биоаккумулятивных поверхностей с инситной органикой.
Калиптровая текстура - достаточно распространенная текстура в рифовых системах фанерозоя. Калип- тра - это органогенная постройка чаще изометрической формы, образованная одним или несколькими организмами, обрастающими друг друга равномерно внутри, либо зонально, либо гнездообразно. Она формируется в активной субплатформенной зоне мелководного шельфа за рифом.
Явные текстуры эрозионной природы [2] наблюдаются в тех участках рифогенных сооружений, которые интенсивно переработаны камнеточцами и илоедами, т. е. подвержены интенсивной биотурбации. Эрогерм- ная текстура представляет собой соотношение биотур- бированной основной массы и частей инситных скелетов (эрокластов), оставшихся после воздействия био- турбаторов. Эроклинная текстура образуется во всех других (кроме биогермных) карбонатных породах, где имеет место активная биотурбационная деятельность грунтоедов, нарушающая первичную дифференциацию карбонатного осадка.
Группа аккумулятивных текстур пород, в которых биокомпоненты являются основными, состоит из двух текстур: клинокластовая и механокластическая - гравитационная. Первая характеризуется соотношением клинокластов и илистой основной массы, заполняющей оригинальные иногда узорчатые промежутки между клинокластами. Механокластическая - гравитационная текстура образуется в тех областях рифового плато, либо зарифовой территории, где механический разнос биокомпонентов и фактор гравитационного осаждения частиц из водной среды уже начинает доминировать.
Биофация - комплекс палеонтологических, биологических или биогенных признаков, характеризующий ту или иную фациальную обстановку. В определение последней входит и географическая, и энергетическая характеристика, особенности рельефа дна, коадаптив- ные возможности организмов. Тип расселения - это жизненная форма сообщества организмов, приспособившихся друг к другу и выработавших общие морфологические черты.
Сообщества организмов вырабатывают общие адаптации как друг к другу, так и к среде, формируя коа- даптации или жизненные формы - типы расселений организмов. Последние «противостоят» следующим факторам: 1) объем поступающего в экологическую нишу осадка; 2) неорганогенная цементация (посредством метеорных вод); 3) растворение метеорными водами; 4) волновая абразия и эрозия; 5) перемывания и перекатывания течениями поверхностного и донного типа; 6) прижизненная органогенная цементация, выраженная в связывании и обрастании скелетов; 7) затвердевание пелитового и илистого материала (субстрата) посредством аккреции; 8) обильное осаждение ила из карбонатонасыщенной среды, инициированное бактериями и микроводорослями.
По области жизнедеятельности среди биофаций [2] можно выделить три группы: бентосные, связанные с жизнью на поверхности субстрата; инфаунные - живущие внутри субстрата и нектонно-планотонные - живущие в толще воды.
Определение состава, типа биофаций и типа расселения организмов для инситных захоронений и захоронений на месте жизнедеятельности (1 и 2 группы) производится с высокой долей интерпретации реально существующих прижизненных соотношений организмов, условий их существования, динамики среды. Среди бентосных биофаций выделяется две группы: жесткоприкрепленные к грунту организмы и свободно-лежащие на грунте (фарины). Фа- рины относятся к достаточно распространенному характеру расселения организмов на грунте, там где отсутствует активная волновая или течениевая деятельность, где достаточно пищи и кислорода, где спокойная не застойная обстановка и небольшой привнос осадка извне.
Аэдисами автором названа группа инфаунных биофаций, в составе которой преобладает ихнофауна, ведущая как активный, так и пассивный образ жизни, как образующие самостоятельный скелет, так и мягкотелые, как формирующие трубки, норы, раковины внутри грунта, так и просто оставляющие следы своей деятельности,
Среди прикрепленных бентосных биофаций установлено четыре типа расселения организмов: луга, поля, тампы и заросли.
Биофация с доминированием стабилизаторов коркового типа или организмов с преобладающей корковой, пленочной, пластинчатой и т. п. формой скелета диагностируется чаще в гидродинамически активной зоне (при скальном субстрате) либо, наоборот, в затишных обстановках (илистый субстрат) и в энергичных (зернисто-пелитовый субстрат) карбонатных платформах. Такой тип расселения организмов, названный «поля», характерен для многих таксономических групп, включая даже бесскелетные формы [2].
Луга - тип расселения организмов с массивной, изометрической или близкой к ним формой скелетов. Это основной и главный для современных и древних погребенных рифов тип расселения организмов-архитекторов, рифостроителей. Функция «конструирования» заложена в генофонде различных таксономических групп: от фо- раминифер до водорослей. Биогенные структуры этого типа расселения обычно представлены инфлаутстоуна- ми и фреймстоунами, а текстуры, в основном массивные, - однородные, биоритмитные, эрогермные.
Заросли - тип расселения организмов с кустистыми, ветвистыми, пластинчатыми, прямостоящими, одиночными, цилиндрическими и т. п. формами скелетов. Главными биогенными структурами «инситных» зарослей являются флаутстоуны и бафлстоуны. Причем структуроопределяющими компонентами в них являлись организмы со скелетом не только рамозного типа, но и с сегментным, спикуловым и, возможно, камерным типом.
Тампы - тип расселения организмов с мелкими желваковыми, изометрическими, грушевидными, якорными формами скелета. Наиболее типичными биогенными структурами для тампов в литифицированном состоянии являются инфлаутстоун, фреймстоун, лайт- стоун, баундстоун, а текстурами - массивные, однородные, биоритмитные, эрогермные.
В классификации первичных биогенных структур (таблица) карбонатных пород автором заложен принцип энергетики среды. Главным в классификации является соотношение илистого и зернистого компонентов. Чем больше ила в осадке, тем ниже энергия среды; чем больше зернистого компонента, тем выше ЭСБ. Это отражено в последовательностях различных типов биокластитов: от мадстоуна до грейнстоуна. Уровни ЭСБ достаточно условные, в настоящей работе выбрано 5 уровней, самый низкий - это первый, а самый высокий - это пятый уровень.
Плотность захоронения, размерность биокомпонентов, а также тип скелетов - важнейшие характеристики, которые могут быть использованы при определении динамики среды. Типизация скелетов по степени их стойчивости к разрушению (классификация R.N. Gins- burq) - главный инструмент при определении компонентов, слагающих основные породообразующие структуры. В ряду биокластитов в обстановке V уровня ЭСБ [2] не может литифицироваться обломочный материал, т.к. здесь доминирует процесс разрушения, а не накопления. Размерность компонентов биогенных структур, как и плотность их литификации в ориктоце- нозах, имеет огромное значение при биоседиментоло- гическом анализе. Среди биокластитов в четвертой энергетической зоне могут литифицироваться рудсто- уны и грейнстоуны. Увеличение плотности захоронения биокомпонентов совместно с увеличением их размеров для рудстоунов является гарантией их литифи- кации. Для биокластитов дальность транспортировки прямо увязывается с уменьшением размеров обломочных частиц.
Генетическая классификация первичных карбонатных пород
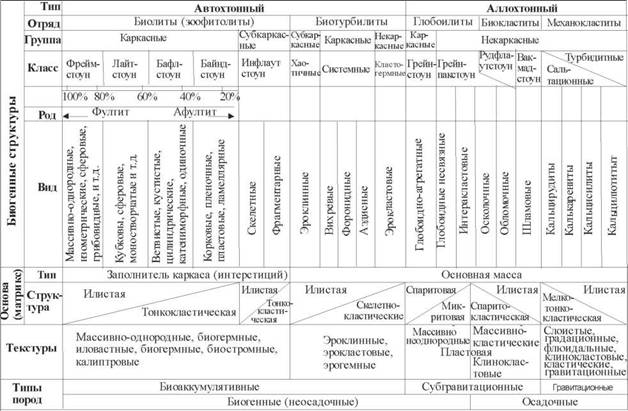
Для биолитов увеличение степени устойчивости скелетов четко коррелирует с увеличением процента содержания автохтона и повышением энергетики среды. Обратная тенденция приводит к образованию (например, в I энергетической зоне) таких биогенных структур, как тонковетвистый бафлстоун, фрагментарный инфлаутстоун. Наличие автохтонных непереме- щенных частиц скелета (например, спикулы губок) - это критерий неподвижной среды.
Третий энергетический уровень, как и все предыдущие, условен и выделяется по сопоставлению биогенных структур в ряду бафлстоунов до инфлаутсто- унов для биолитов, и в ряду от вакстоунов до грейн- стоунов - для биокластитов. Присутствие спарита в пакстоунах в любом количестве, а также в различных разновидностях флаутстоунов - явный показатель повышения ЭСБ.
В ряду биоаккумулятивных образований III энергетический уровень диагностируется в последовательностях биогенных структур от фрагментарного инфлаут- стоуна до викстоуна и софтстоуна. Здесь основными признаками для определения уровня ЭСБ является плотность захоронения и размерность биокомпонентов. Увеличение плотности для этих структур четко коррелирует с увеличением активности среды, одновременно с уменьшением размерности компонентов. Последнее типично только для третьего уровня ЭСБ, в целом же с первого по пятый уровни отмечена прямая зависимость размера скелетов и энергетики среды. Вик- и софтсто- уны насыщены изолированными друг от друга мелкими самостоятельными органогенными постройками до симбиогермов, калиптров и биогермов. Они составляют тоже жесткую систему, которую часто называют субкаркасной, т.к. она является достаточно устойчивой к волновому воздействию. Второй энергетический уровень характеризуется в системе биолитов наличием флаутстоунов и фрагментарных инфлаутстоунов ветвистого и камерного типов.
Большинство современных классификаций карбонатных пород основано на тех же принципах, что и классификации терригенных. Главным из них является принцип дальности переноса материала, коррелируемый с размерами обломочных частиц.
В классификациях осадочных пород, содержащих органические остатки, в советской литературе, начиная с начала 1950-х гг. [8-15] используется только таксономический состав породообразующих организмов. Много терминов в старых систематиках, которые имеют в словосочетаниях противоречия, например органогенно-обломочные, биоморфно-детритовые, раковинно-детритовые, водорослево-биогермные. Из всех советских классификаций заслуживает внимания классификация Г.И. Теодоровича в 1950-1958 гг. [9], в которой реализовалась идея автохтонных неперемещенных карбонатных образований и пород аллохтонных, т. е. явно обломочного характера.
М.С. Швецов [8] установил 4 класса: обломочных пород (с подклассами терригенно-обломочных и пи- рокластических); химических - биохимических; кау- стобиолитов и смешанных пород. Карбонатные породы по этой классификации попадают в группу биохимических пород с участием организмов. Это не в полной мере отражает генетическую природу и разнообразие карбонатных пород и образований.
Первичные карбонатные породы ряда кальцит- арагонит подразделяются на две большие группы (см. таблицу): биогенные и осадочные. Первая группа характеризуется доминированием биогенного фактора в образовании структур и текстур и в преобладании биокомпонентов в составе. Вторая группа объединяет первичные карбонатные породы типично осадочного происхождения, т. е. в процессе образования которых есть фактор механического или физического разрушения литифицированных образований, фактор водного переноса и фактор осаждения перенесенных частиц. В группу настоящих осадочных пород не входит отряд глобоилитов - аллохтонных по сути, но не типично осадочных, т. к. для этого отряда не характерны первый и последний факторы осадочного процесса.
Гравитационные первичные осадочные карбонатные породы (механокластиты) подразделяются на два класса: сальтационные - с механической обработкой зерен и турбидитные, или потоковые. По размеру компонентов среди механокластитов выделяются четыре вида пород: кальцирудиты, калькарениты, кальцисил- титы и кальцилюлиты.
В понимании генезиса карбонатных пород важнейшее значение имеет представление об «основе» или матриксе. Для аллохтонных субгравитационных и гравитационных осадочных пород в качестве матрикса служит основная масса, состав которой меняется от илистого до тонкомелкокластического в зависимости от динамики разрушения пород, скорости и дальности переноса и условий садки материала.
Отряд аллохтонных (перемещенных) первичных осадочных пород - биокластитов - разделяется на шесть классов структур: грейнстоун, пакстоун, рудсто- ун, флаутстоун, вакстоун и мадстоун в полном соответствии с классификацией Р. Данхэма [4], А. Эмбри и Дж. Кловена [5].
Пропорции ила, спарита и мелкой кластики в основной массе в соотношении с упаковкой главных структурообразующих компонентов четко отражают динамическое состояние среды седиментации. По доминирующему составу компонентов биокластиты разделяются на три вида: осколочные, обломочные и шламовые в зависимости от степени разрушения, окатан- ности, физического истирания компонентов и наличия тонкого карбонатного ила как продукта их разрушения. Отсутствие ила и присутствие спарита - показатели высокого динамического режима седиментации.
В составе глобоилитов два рода биогенных структур: грейпстоун (грейпстоун - это связный грейнстоун) и пакстоун. Они отличаются от биокластических грейн-пакстоунов только доминантным составом структурообразующих компонентов. Глобоилиты не относятся к типичным осадочным породам, а представляют собой биогенные образования с элементом биоаккумуляции.
Типичные автохтонные биогенные первичные карбонатные породы представляют собой не осадочные, а биоаккумулятивные образования. В их составе выделяются два отряда: биотурбилиты и биолиты (с подотрядами: зоолиты и фитолиты). Биотурбилиты - новая группа пород, объединяющая все породы, возникшие в результате активного воздействия (эродирующего, биотурбационного) организмов на субстрат как каменистый, ранее литифицированный, так и синхронный времени биоаккумуляции.
В составе биотурбилитов выделяют три группы образований: каркасные, субкаркасные и некаркасные. К первому отнесены системные биотурбилиты, которые в плотных популяциях образуют связные между собой постройки биотурбаторов, имеющие свою собственную внешнюю скелетную оболочку. К субкаркасным био- турбилитам отнесены эроклинные разности, в которых в качестве «каркаса» можно рассматривать систему биологически переработанных поверхностей. Эрокла- стовые биотурбилиты не имеют каркаса, т. к. представляют собой чаще «песчаники», сложенные эрокласти- ческим материалом (рис. 1).
Автохтонные биолиты - это главные биоаккумуля- тивные биогенные породы. К осадочным породам их относить нельзя, т. к. типичного осадка в них мало, а влияние гравитации выражается чаще в направлении роста породообразующих организмов и в меньшей степени в дифференциации («слоистости») илистого материала в заполнителе каркаса. Матрикс этих пород следует называть заполнителем интерстиций между скелетами, а не основной массой как для субгравитационных и гравитационных карбонатолитов. Состав заполнителя интерстиций меняется от илистого до тонкокластиче- ского в зависимости от гидродинамической обстановки. Причем отсутствие кластики и ила в интерстициях каркаса может интерпретироваться равнозначно как признак среды с наивысшей гидродинамической активностью.
В составе отряда биолитов два подотряда: зоолиты и фитолиты, различающиеся тем, что в первом доминируют животные, а во втором - растительные сообщества породообразующих организмов. В отряде биолитов две группы биоаккумулятивных образований: субкаркасные и каркасные. Субкаркасные биолиты характеризуются биогенной структурой: инфлаутстоун. Плотность захоронения скелетов в инфлаутстоуне, их размеры, соотношение с количеством интерстиций и характером (ил, спарит, тонкая кластика) заполнителя интерстиций - это признаки, по которым можно судить о повышении либо понижении гидродинамического состояния режима биоаккумуляции. В составе инфла- утстоунов два вида образований: скелетные и фрагментарные.
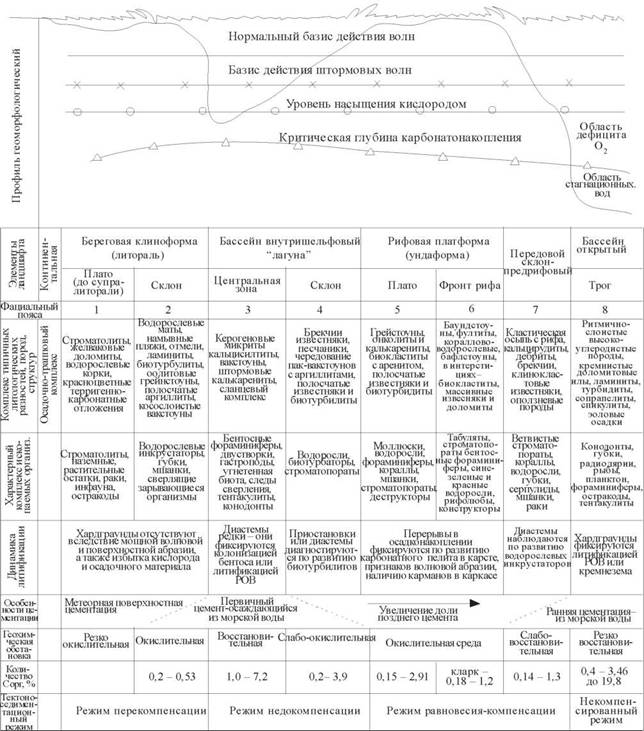
Рис. 1. Идеализированный фациальный профиль морских обстановок в силуре - девоне
Каркасные биолиты разделяются на четыре класса: фреймстоун, байндстоун, бафлстоун и лайтстоун. Первые три установлены еще А. Эмбри и И. Кловэном в 1971 г. [5], а в отечественной литературе описаны в работе «Современные и ископаемые рифы. Термины и определения» в 1990 г. [16], а также в последних классификациях В.Н. Шванова, В.Т. Фролова, Э.И. Сергеевой и др. в 1998 г. [14]. Новым подразделением в классе каркасных биолитов является лайтстоун, оха- растеризованный в 1986 г. Г. Д. Исаевым [7]. Он представляет собой каркасное сооружение, образованное организмами с камерным типом скелета. К таковым можно отнести некоторых брахиопод, фораминифер, устриц, рудистов и т. п.
Три класса основных каркасных биолитов отличаются доминированием структуроопределяющего биокомпонента: для фреймстоуна характерно преобладание скелетов массивного типа, для баундстоунов - коркового, а для бафлстоунов - ветвистого или рамозного типа. Разновидности или виды отдельных категорий каркасных биолитов определяются (см. таблицу) разнообразием морфологии скелетов организмов, преобладающих в данной популяции. Особую разновидность следует выделить в составе фреймстоунов-байнд- стоунов - образование, сложенное на 100% из скелетов массивного либо коркового типа, т. е. почти без заполнителя и пустот между скелетами. Ранее автор называл их биолититами по Р. Фолку [17]. Однако, учитывая неопределенность этого термина, заложенную его автором, вместо биолитита здесь предлагается новый термин: фултит (от англ. full - полный). Другую крайнюю разновидность каркасных биолитов с максимальным количеством заполнителя каркаса (например, для пленочных байдстоунов - до 90% объема породы) и минимальным объемом каркасообразующего биокомпонента в породе следует называть афултит, т.е. неполный.
Размер породообразующих компонентов в каркасных биолитах, плотность их упаковки в породе (плотная, насыщенная, ненасыщенная или неплотная), соотношение ила, спарита и кластического материала в заполнителе каркаса и количество последнего - это признаки, по которым можно судить об уровне гидродинамического состояния бассейна биоаккумуляции. Распределяя вышеназванные признаки в определенной иерархии мы создаем конкретный инструмент для определения фациального типа карбонатных пород. Таким образом, настоящая схема генетической классификации первичных карбонатных пород является своеобразной основой регионального фациального анализа в областях карбонатонакопления (см. рис. 1).
Осадочные породы, доминирующие в верхних горизонтах литосферы, являются главными носителями полезных ископаемых. Предлагаемая в настоящей работе авторская схема классификации карбонатных пород (см. таблицу) отличается от ранее известных следующими признаками.
Схема классификации посвящена только первичным (а не преобразованным) карбонатным породам кальцит-арагонитового ряда.
Схема основана не на структурно-вещественном составе пород, а на текстурно-компонентно-структурном строении первичной карбонатной породы.
В основу схемы положены биоседиментологиче- ские признаки, роль организмов в образовании пород и воздействии их на субстрат, а также гидродинамические признаки среды биоаккумуляции.
Схема составлена на базе обширных знаний палеонтологии породообразующих организмов, их секре- ционной деятельности, геометрии и стереометрии ске- летообразования. Незнание этой сферы обычно приводит к выделению таких категорий, как «органогеннообломочные», «биогенные с реликтами теней» и т.д.
Классификация первичных карбонатных пород в основе своей опирается на систему структур зарубежных авторов: Р. Данхема (1962), А. Эмбри и Дж. Кло- вена (1971). Именно эта система является единственной, которая построена на жизненных формах популяций породообразующих организмов.
В настоящей схеме при определении компонентного состава отдельных категорий первичных пород используется классификация скелетов (Гинсбург, 1956) по степени их устойчивости к разрушению.
Таким образом, используя виды, классы и группы структур и типов пород генетической классификации (см. таблицу) совместно с другими признаками идиали- зированного профиля (см. рис. 1), можно достаточно надежно определить фациальную закономерность (пояса с 1-го по 8-й) силурийских и девонских бассейнов практически в пределах всей Западно-Сибирской плиты.
Список литературы
Друшиц В.В. Палеонтология беспозвоночных. М.: МГУ, 1974. 528 с.
Исаев Г.Д. Основы биоседиментологии и региональный фациальный анализ. Новосибирск: Гео, 2006. 132 с.
Ginsburg R.N. Environmental relatinships of grain size and constituent particles in some south Florida carbonate sediments // Bull. Amer. Assoc. Pet rol. Geol. 1956. Vol. 40, № 10. P. 2384-2427.
Danham R.J. Classification of carbonate rocks according to depositional texture Classification of carbonate rocks: Simposium Amer. Assoc. Petrol.
Geol. Mem / Ed. by V.E. Ham. 1962. Vol. 1. P. 108-121.
Embry A.F., Klovan J.E. A late devonian reef tract on northheastehn Bancks Jsland Northwest Territories // Bull. Can. Petrol. Geol. 1971. Vol. 19, № 4. P. 730-781.
УилсонДж.Л. Карбонатные фации в геологической истории. М.: Недра, 1980. 464 с.
Исаев Г.Д. Методика изучения биогенных структур карбонатных пород палеозоя для целей усовершенствования региональной стратиграфи ческой основы // Палеоэкологический и литолого-фациальный анализы для обоснования детальности региональных стратиграфических схем. Новосибирск: СНИИГИМС, 1986а. С. 91-100.
ШвецовМ.С. Петрография осадочных пород. М.: Госгеолтехиздат, 1958. 416 с.
Теодорович Г.И. О минералого-геохимических особенностях первичных нефтеносных и рассеянно-битуминозных свит // Минералогия и
фации битуминозных свит ряда областей СССР. М.: АН СССР, 1962. С. 61-77.
ХвороваИ.В. Атлас карбонатных пород среднего и верхнего девона Русской платформы. М.: АН СССР, 1958. 170 с.
Казанский Ю.П., Белоусов А.Ф., Петров В.Г. и др. Осадочные породы (классификация, характеристика, генезис). Новосибирск: Наука, 216 с.
Фролов В.Т. Основы генетической типизации морских отложений // Мор. геол. седиментол., осад. петрогр. и геол. океана. Л., 1980. С. 4247.
Шванов В.Н. К методике описания осадочных формаций по структурно-вещественным признакам // Известия вузов. Геология и разведка. № 4. С. 3-9.
Систематика и классификация осадочных пород и их аналогов / В.Н. Шванов, В.Т. Фролов, Э.И. Сергеева и др. СПб.: Недра, 1998. 352 с.
Тихомиров С.В. Вопросы генетической классификации осадочных пород // Бюлл. МОИП. Отд. геол. 1986. Т. 61, № 2. С. 140-141.
Современные и ископаемые рифы. Термины и определения: Справочник / И.Т. Журавлева, В.Н. Космынин, В.Г. Кузнецов и др. М.: Недра, 1990. 184 с.