| Скачать .docx | Скачать .pdf |
Реферат: Структура и свойства биологических мембран. Транспорт вещств в биологических мембранах
Структура и свойства биологических мембран. Транспорт вещств в биологических мембранах
Биологическая мембрана - это структура, состоящая из органических молекул, которая имеет толщину около 7-10нм и видима только посредством электронного микроскопа. В каждой клетке есть плазматическая мембрана, которая ограничивает содержимое клетки от наружней среды, и внутренние мембраны, которые формируют различные органоиды клетки (митохондрии, органоиды, лизосомы и т.п.)
Плазматическая мембрана выполняет несколько важных функций.
1) Образует избирательный барьер, который отделяет содержимое клетки от окружающей среды, что позволяет поддерживать постоянными химический состав цитоплазмы и её физические свойства.
2) Регулирует транспорт веществ между содержимым клетки и окружающим клетку раствором.
3) Принимает участие в информационных процессах в живой клетке.
Химическая состав и структура плазматической мембраны
В состав плазматической мембраны входят липиды, белки и углеводы. Соотношение между липидами и белками может значительно варьировать в различных клетках.
Липиды мембраны бывают трех видов: глицерофосфолипиды, сфингофосфолипиды и стероиды (холестерол).
Молекула глицерофосфолипида состоит из остатка трёхатомного спирта глицерола, атомы водорода двух гидроксильных групп которого замещены на две длинные цепи жирных кислот. Третий атом водорода гидроксильной группы глицерина замещён остатком фосфорной кислоты, к которому, в свою очередь, присоединён остаток одного из азотистых оснований (холин, этаноламин, серин, инозитол).
В молекуле глицерофосфолипида можно выделить две части, которые называются головка (остаток глицерина, остаток фосфорной кислоты и азотистое основание) и хвостики (остатки жирных кислот). Головка и хвостики сильно отличаются по своим физическим свойствам. Головка молекулы фосфолипида гидрофильна (″любит воду″). Она хорошо растворима в воде. Хвостики - гидрофобны (″боятся воды″). Они легко растворяются в липидах и органических растворителях, но водой отталкиваются. Таким образом, в целом молекула фосфолипида, содержащая как водорастворимые, так и липидорастворимые области, имеет амфифильные свойства.
Молекулы сфингофосфолипидов также состоят из головки и хвостиков. Они отличаются из фосфолипидов тем, что вместо остатка глицерина содержат остаток спирта сфингозина.
Если сухие фосфолипиды погружают в воду, они спонтанно формируют в зависимости от их концентрации различные структуры (Рис. 1). Одна из них - сферическая структура, называемая мицеллой. Молекулы фосфолипидов упорядочены так, что гидрофильные головки направлены в водную среду, а гидрофобные хвосты - внутрь структуры.
При более высокой концентрации фосфолипидов, их молекулы формируют бислойные пластинчатые структуры. Немецкие ученые Gorter и Grendel доказали, что такая бислойная фосфолипидная структура является основой мембраны клетки.
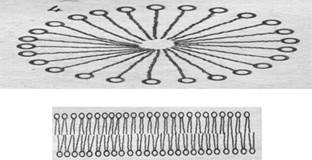
Рис. 1. Мицелла и бислойная пластина в водном растворе
Физическое состояние фосфолипидного бислоя зависит от температуры. Если температура превышает критическую точку, бислой представляет собой жидкость. При этом каждая молекула имеют возможность перемещаться.
Существует несколько видов движения молекул липидов: колебание, вращение, латеральная диффузия (перемещение молекул в пределах своего слоя), флип-флоп (перемещение молекул из одного слоя липидов в другой, происходит редко).
Если температура падает ниже критической точки, мембранные фосфолипиды становятся твердыми. Мембрана теряет текучесть, и движение молекул в ней ограничивается.
Согласно современной жидкостно-мозаичной модели мембраны (модель Сингера и Николсона), липидный бислой является основой мембраны. Молекулы фосфолипидов расположены в нём так, что их длинные оси параллельны и ориентированы перпендикулярно к поверхности мембраны. Мембрана сохраняется в жидком состоянии благодаря температуре клетки и химическому составу жирных кислот.
Белки мембраны подразделены на два вида. Молекулы первого типа являются гидрофильными. Эти белки, называемые периферическими, соединены с поверхностью мембраны сравнительно слабыми электростатическими силами. Белки второго вида имеют как гидрофильные, так и гидрофобные группы. Их молекулы более или менее погружены в мембрану, и удерживаются в ней более прочными гидрофобными силами. Некоторые белки пронизывают мембрану от ёё внутренней до внешней поверхностей - интегральные белки (Рис. 2).
Многочисленные белки мембраны выполняют различные функции (метаболическую, транспортную, рецепторную и т.п.). Функции белков мембраны существенно зависят от строения их молекул.
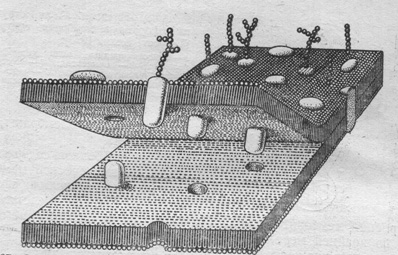
Рис. 2. Жидкостно-мозаичная модель мембраны: фосфолипидный бислой; периферические и интегральные белки.
Классификация процессов транспорта в биологических мембранах
Мембрана клетки является избирательным барьером для различных веществ, находящихся внутри и снаружи клетки. Существует несколько специфических механизмов транспорта в мембранах. Все он могут быть подразделены на два типа: пассивный и активный транспорт.
Все виды пассивного транспорта основаны на принципе диффузии. Небольшая частица, растворённая в жидкости, постоянно подвергается ударам со стороны окружающих её молекул жидкости. Результатом этого является хаотическое движение частицы, которое называется броуновским движением. Диффузия является результатом хаотических независимых движений многих частиц. Если концентрация вещества одинаковая в каждой части раствора, то движение частиц хаотично. При этом существует дрейф частиц из областей, где они расположены более плотно, в области, где частиц меньше.
Диффузия незаряженных частиц вызывается их концентрационным градиентом и направлена в сторону уменьшения этого градиента. Частицы вещества перемещаются из области более высокой концентрации вещества в области, где концентрация этого вещества низкая. Диффузия постепенно уменьшает градиент концентрации до тех пор, пока не наступит состояние равновесия. При этом в каждой точке установится равная концентрация, и диффузия в обоих направлениях будет осуществляться в равной степени. Диффузия является пассивным транспортом, поскольку не требует затрат внешней энергии.
Существует несколько видов диффузии в плазматической мембране:
1) Свободная диффузия.
2) Облегченная диффузия неэлектролитов.
3) Электродиффузия (облегченная диффузия ионов).
Раствор вещества высокой концентрации обладает более высокой свободной энергией, чем раствор вещества более низкой концентрации. В процессе диффузии энергия рассеивается. Напротив, вещество не может переместиться из области низкой его концентрации в область высокой его концентрации за счёт внутренней энергии. Для этого необходима дополнительная энергия из внешнего источника.
Для того, чтобы перемещать вещества против их концентрационного или электрохимического градиентов, мембрана использует энергию метаболизма. Такой тип транспорта называется активным транспортом. Есть два основных вида активного транспорта:
1) Первично-активный транспорт.
2) Вторично-активный транспорт.
Более сложные механизмы транспорта - экзоцитоз и эндоцитоз, в ходе которых макромолекулы поступают в клетку или выделяются из неё через небольшие, окружённые мембраной везикулы.
Свободная диффузия
Вещества, перемещающиеся через мембрану путём свободной диффузии, не образуют каких-либо химических связей с другими веществами.
Для количественной характеристики диффузии используют физическую величину - поток вещества (J) : J = dn/dt · 1/S (1),
где n - количество вещества в молях, перемещающихся посредством диффузии через поверхность S, перпендикулярную потоку вещества, за единицу времени.
Первый закон Фика указывает, что поток вещества, перемещаемого путём диффузии, пропорционален движущей силе диффузии - градиенту концентрации вещества: J = - D · dC/dx (2).
Отрицательный знак означает, что поток направлен из области высокой концентрации вещества в область с более его низкой концентрацией, в результате чего градиент концентрации уменьшается. D - коэффициент диффузии, который зависит от природы вещества и температуры:
D = U·R·T (3),
где U - подвижность частиц вещества, R - универсальная газовая постоянная, T - абсолютная температура.
Если диффузия осуществляется через мембрану, уравнение (2) может быть представлено как J = -P · (C1 - C2) (4),
где C1 и C2 - концентрация раствора внутри и вне клетки, P - коэффициент проницаемости мембраны для данного вещества. Коэффициент проницаемости определяется коэффициентом диффузии D вещества, толщиной мембраны d и коэффициентом распределения вещества K, зависящим от растворимости вещества в органических растворителях, но не воде.
P = Dk/d (5)
Проницаемость мембраны для неэлектролитов существенно зависит от их способности растворяться в билипидном слое мембраны. Проницаемость мембраны для различных веществ определяют по растворимости в оливковом масле, которую можно рассматривать как модель мембранных липидов. Таким образом, мембрана хорошо проницаема для липидорастворимых веществ (спирты, эфиры), не имеющих биологического значения. Но такие гидрофильные вещества как сахара, аминокислоты не способны проникать через биологическую мембрану посредством свободной диффузии. Для этого требуются специальные системы транспорта (смотри ниже).
Проницаемость мембраны зависит также от размера молекул. Мелкие молекулы могут проникать через мембрану путём свободной диффузии. Например, вода не растворима в липидах и органических растворителях. Но она проникает через плазматическую мембрану благодаря небольшому размеру молекул. Проницаемость мембраны для воды очень высокая. Предполагают, что она проникает в мембрану через временные структурные дефекты, формирующихся при тепловых колебаниях хвостиков из жирных кислот. Эти дефекты (кинки) позволяют перемещаться через мембрану не только молекулам воды, но также другим небольшим гидрофильным молекулам (кислород, углекислый газ).
Облегченная диффузия
Крупные гидрофильные молекулы (сахара, аминокислоты) перемещаются через мембраны с помощью специальных молекул - мембранных переносчиков. Мембранные переносчики представляют собой интегральные белки, которые имеют центры связывания транспортируемых молекул. Образующаяся связь белка и переносчика является обратимой и обладает высокой степенью специфичности. Транспортируемая молекула проходит через мембрану вследствие изменения конформации белка-переносчика при химическом взаимодействии центров связывания обеих молекул.
Транспорт веществ через мембрану, в котором используются транспортные молекулы, называются облегчённой диффузией. Этот тип транспорта мембраны является одним из видов диффузии, поскольку транспортируемое вещество перемещается по градиенту концентрации. Никакая дополнительная энергия не требуется для этого процесса. Но облегченная диффузия отличается от свободной диффузии своей высокой специфичностью. Переносчики мембраны могут узнавать даже оптические изомеры одного и того же вещества.
Другой особенностью облегченной диффузии является феномен насыщения. Поток вещества, транспортируемого путём облегченной диффузии, растёт в зависимости от концентрации вещества только до определенной величины. Затем возрастание потока прекращается, поскольку транспортная система полностью занята. Таким образом, действие транспортной системы подобное катализу ферментами, однако переносчик не ускоряет химическую реакцию, а перемещает вещество через мембрану.
Существуют некоторые системы переносчиков, которые способны транспортировать более одного вещества. Процесс называется симпортом (или котранспортом), если вещества перемещаются в одном и том же направлении, и антипортом (встречным транспортом), если направления перемещения веществ противоположны.
Примером облегченной диффузии является действие системы транспорта глюкозы через мембраны эритроцитов и мышечных клеток. Другой пример - антипорт бикарбоната и ионов гидроксила в плазматической мембране эритроцитов.
Электродиффузия
Электродиффузия - диффузия электрически заряженных частиц (ионов) под влиянием концентрационных и электрических градиентов. Ионы - атомы или группы атомов, которые приобретают электрический заряд, теряя или приобретая электроны. Липидный бислой мембраны непроницаем для ионов. Они могут проникнуть через плазматическую мембрану только посредством специальных структур - ионных каналов, которые образованы интегральными белками.
Движущей силой диффузии является не только разность концентрации ионов внутри и вне клетки, но также разность электрических потенциалов, создаваемых этими ионами по обе стороны мембраны. Следовательно, диффузионный поток ионов определяется градиентом электрохимического потенциала (электрохимический градиент).
Электрохимический потенциал является энергией ионов:
|
|
μ0- стандартный химический потенциал, который зависит от химической природы вещества и температуры, R - универсальная газовая постоянная, T - температура, C - концентрация иона, z - электрический заряд, F - константа Фарадея, φ - электрический потенциал.
Зависимость потока ионов J от электрохимического градиента определяется уравнением Теорелла:
|
|
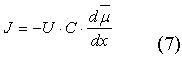
где U - подвижность ионов, C - концентрация ионов, dμ/dx - электрохимический градиент.
Подставляя (6) в (7), можно получить уравнение Нернста-Планка с учётом двух градиентов, которые обуславливают диффузию ионов:
|
|
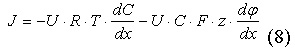
Ионные каналы мембраны представляют собой интегральные белки мембраны, которые образуют отверстия в мембране, заполненные водой. В плазматической мембране обнаружен ряд ионных каналов, которые характеризуются высокой специфичностью, допускающей перемещение только одного вида ионов. Существуют натриевые, калиевые, кальциевые и хлорные каналы. Каждый из них имеет так называемый селективный фильтр, который способен пропускать только определённые ионы. Существует несколько теорий, объясняющих избирательность ионных каналов плазматической мембраны.
Проницаемость ионных каналов может изменяться благодаря наличию ворот, определенных групп атомов в составе белков, формирующих канал. Конформационные изменения ворот переводят канал из открытого состояния в закрытое и наоборот. Механизмы регуляции положения ворот могут отличаться в различных каналах. Некоторые из них открываются при изменениях электрического потенциала мембраны. Другие открываются под действием специфических химических веществ, выполняющих сигнальные функции.
Первично-активный транспорт
Действие пассивного транспорта через мембрану, в ходе которого ионы перемещаются по их электрохимическому градиенту, должно быть сбалансировано их активным транспортом против соответствующих градиентов. В противном случае, ионные градиенты исчезли бы полностью, и концентрации ионов по обе стороны мембраны пришли бы в равновесие. Это действительно происходит, когда активный транспорт через мембрану блокируют охлаждением или путём использования некоторых ядов.
Существует несколько систем активного транспорта ионов в плазматической мембране (ионные насосы):
1) Натрий-калиевый насос.
2) Кальциевый насос.
3) Водородный насос.
Активный транспорт - перенос ионов против их электрохимических градиентов с использованием энергии метаболизма:
Натрий-калиевый насос существует в плазматических мембранах всех животных и растительных клеток. Он выкачивает ионы натрия из клеток и загнетает в клетки ионы калия. В результате концентрация калия в клетках существенно превышает концентрацию ионов натрия.
Натрий-калиевый насос - один из интегральных белков мембраны. Он обладает энзимными свойствами и способен гидролизовать аденозинтрифосфорную кислоту (АТФ), являющуюся основным источником и хранилищем энергии метаболизма в клетке. Благодаря этому указанный интегральный белок называется натрий-калийиевой АТФазой. Молекула ATФ распадается на молекулу аденозиндифосфорной кислоты (АДФ) и неорганический фосфат.
Таким образом, натрий-калиевый насос выполняет трансмембранный антипорт ионов натрия и калия. Молекула насоса существует в двух основных конформациях, взаимное преобразование которых стимулируется гидролизом ATФ. Эти конформации выполняют функции переносчиков натрия и калия. При расщеплении натрий-калиевой АТФазой молекулы ATФ, неорганический фосфат присоединяется к белку. В этом состоянии натрий-калиевая АТФаза связывает три иона натрия, которые выкачиваются из клетки. Затем молекула неорганического фосфата отсоединяется от насоса-белка, и насос превращается в переносчик калия. В результате два иона калия попадают в клетку. Таким образом, при расщеплении каждой молекулы ATФ, выкачиваются три иона натрия из клетки и два иона калия закачиваются в клетку. Один натрий-калиевый насос может перенести через мембрану 150- 600 ионов натрия в секунду. Следствием его работы является поддержание трансмембранных градиентов натрия и калия.
Через мембраны некоторых клеток животного (например, мышечных) осуществляется первично-активный транспорт ионов кальция из клетки (кальциевый насос), что приводит к наличию трансмембранного градиента указанных ионов.
Водородный ионный насос действует в мембране бактериальных клеток и в митохондриях, а также в клетках желудка, перемещающего водородные ионы из крови в его полость.
Вторично-активный транспорт
Существуют системы транспорта через мембраны, которые переносят вещества из области их низкой концентрации в область высокой концентрации без непосредственного расхода энергии метаболизма клетки (как в случае первично-активного транспорта). Такой вид транспорта называется вторично- активным транспортом.
Вторично-активный транспорт некоторого вещества возможен только тогда, когда он связан с транспортом другого вещества по его концентрационному или электрохимическому градиенту. Это симпортный или антипортный перенос веществ.
При симпорте двух веществ ион и другая молекула (или ион) связываются одновременно с одним переносчиком прежде, чем произойдёт конформационное изменение этого переносчика. Так как ведущее вещество перемещается по градиенту концентрации или электрохимическому градиенту, управляемое вещество вынуждено перемещаться против своего градиента.
Ионы натрия являются обычно ведущими веществами в системах симпорта клеток животного. Высокий электрохимический градиент этих ионов создаётся натрий-калиевым насосом. Управляемыми веществами являются сахара, аминокислоты и некоторые другие ионы. Например, при всасывании питательных веществ в желудочно-кишечном тракте глюкоза и аминокислоты поступают из клеток тонкой кишки в кровь путём симпорта с ионами натрия. После фильтрации первичной мочи в почечных гломерулах, эти вещества возвращаются в кровь той же системой вторично-активного транспорта.
Эндоцитоз и экзоцитоз
Макромолекулы - белки и нуклеиновые кислоты - не могут проникнуть через плазматическую мембрану с помощью механизмов транспорта, рассмотренных выше, из-за своих больших размеров. При трансмембранном транспорте больших молекул сама плазматическая мембрана подвергается согласованным перемещениям, вследствие которых часть жидкой внеклеточной поглощается (эндоцитоз) или часть внутренней среды клетки выделяется (экзоцитоз).
В процессе эндоцитоза плазматическая мембрана окружает часть внешней среды, формируя вокруг неё оболочку, в результате чего образуется везикула, которая поступает внутрь клетки. При пиноцитозе образуются небольшие, заполненные жидкостью везикулы. В процессе фагоцитоза формируются большие везикулы, которые содержат твердый материал, например, клетки бактерий.
При экзоцитозе транспортируемое вещество синтезируется в клетке, связывается мембраной в везикулы и экспортируется из клетки. Таким образом транспортируются из клетки специфические белки, нуклеиновые кислоты, нейромедиаторы и т.п.