| Скачать .docx | Скачать .pdf |
Доклад: Доклад: Наша история, записанная в ДНК
Платон очень гордился своим определением человека: двуногое без перьев. Оно имело большой успех, пока киник Диоген, ощипав петуха, не изготовил наглядное пособие. Со времен Платона и Диогена появилось немало новых определений, в том числе основанных на описании свойств молекулы ДНК.
Для дальнейшего изложения важно представить себе молекулу ДНК в виде текста, определяющего многие наши особенности как биологического вида. Текст этот записан последовательностью из молекулярных букв — нуклеотидов. В генетическом алфавите их всего четыре: А (аденин), Т (тимин), Г (гуанин) и Ц (цитозин). Общая длина нуклеотидной последовательности, получаемой от каждого из родителей, — 3 млрд букв.
Генетический текст человека немногим отличается от текстов близкородственных видов. Так, в сходных генах человека и шимпанзе обычно из 100 букв текста 99 одинаковы. А у двух человек (если они не однояйцевые близнецы) в среднем из тысячи только один нуклеотид другой. Именно с такими заменами связаны наследуемые индивидуальные особенности каждого человека.
В последние несколько лет изучение разнообразия генетических текстов (геномов) людей стало одной из самых популярных областей науки. Здесь есть чисто практический интерес: ведь с генетическими особенностями связано здоровье человека. Сегодня в геномные исследования фармацевтические компании вкладывают огромные средства и надеются получить отдачу в ближайшие десятилетия в виде принципиально новых методов диагностики и лечения.
Есть и другой аспект таких разработок: они позволяют реконструировать события давнего прошлого, восстановить историю возникновения вида Homo sapiens и пути миграций разных народов. Эти исследования привели к появлению новых направлений науки — молекулярной антропологии и палеогеномики.
Молекулярные часы и филогенетическое древо
Еще в 60-е годы Л.Полинг и Э.Цукеркандл высказали идею, что скорость накопления мутаций достаточно постоянна: ее можно использовать как своего рода молекулярные часы эволюционной истории. Позже, когда появились методы чтения нуклеотидных последовательностей (секвенирования), скорость накопления мутаций была установлена при сравнении ДНК тех видов, время расхождения которых определялось по палеонтологическим находкам.
Метод молекулярных часов приложим и к изучению событий эволюционной истории человека. Здесь удобным инструментом исследования служит митохондриальная ДНК (мтДНК) и Y-хромосома. Остальные хромосомы при передаче потомству обмениваются сходными кусками генетического текста — рекомбинируют. В результате в каждой хромосоме возникают новые, отличные от родительских, комбинации, что существенно затрудняет анализ накопления мутаций.
Y-хромосома передается только по мужской линии — от отца к сыну. Она присутствует в геноме мужчин в единственной копии и потому ни с чем не рекомбинирует (за исключением небольших участков на концах хромосомы, не влияющих на анализ нерекомбинирующей части). Раз появившись, мутации в Y-хромосоме сохраняются на протяжении тысяч поколений, маркируя тем самым всех потомков индивида, у которого они первоначально возникли. Современные мужчины, имеющие одинаковые мутации в Y-хромосоме, восходят к общему предку по мужской линии. Очевидно, что родные братья, получившие от отца Y-хромосомы с одинаковыми текстами, относятся к одной линии. Однако если у одного из братьев произойдет мутация, которую он передаст своим сыновьям, линии наследования разойдутся на две ветви.
Сейчас созданы базы данных, содержащие информацию о Y-хромосомах десятков тысяч людей со всех уголков Земли. Современные методы генетического и статистического анализа таких больших выборок позволяют установить порядок возникновения мутаций и нарисовать филогенетическое древо, описывающее родственные связи всех людей по мужской линии. Чем раньше возникла мутация, тем ближе к корню древа она располагается и тем больше ветвей будут ее содержать. Так как материал, доступный для исследователя, — это ДНК, полученная от наших современников, филогенетическое древо строится от концов ветвей к корню.
При известной скорости накопления мутаций в расчете на поколение, число мутаций, различающих двух людей, можно перевести в абсолютное время, прошедшее с момента появления их генетических линий, т.е. установить, когда жил последний общий предок этих двух людей. При изучении отцовской линии анализируют около полутора-двух десятков изменчивых участков ДНК (SNP- и STR-маркеры) в Y-хромосоме, размер которой составляет 60 тыс. пар оснований.
Эволюционную историю женской линии можно проследить по митохондриальной ДНК (мтДНК). Плод любого пола получает ее только от матери, поскольку в спермии нет митохондрий. В сравнении с ДНК хромосом это очень маленькая молекула — у человека всего 16 500 пар нуклеотидов. В отличие от ядерных генов, большинство которых представлено одной (в Y-хромосоме) или двумя копиями (в остальных хромосомах), в каждой клетке содержится до нескольких десятков тысяч копий молекул мтДНК. Кроме того, эта молекула — кольцевая, а, как известно, такие молекулы стабильнее, чем линейные. Именно эти особенности позволили исследователям прочесть мтДНК из останков древних людей, в том числе и неандертальцев. Можно считать, что митохондриальная ДНК — подарок природы молекулярным биологам.
Для построения филогенетического древа по материнской линии достаточно определить последовательность нуклеотидов небольших (около 400 пар нуклеотидов) фрагментов, которые мутируют наиболее быстро. Другой метод анализа — определение изменений в участках ДНК, распознаваемых ферментами, рестриктазами. Для большей точности используют комбинацию обоих методов. Поскольку технически изучать мтДНК проще, такие работы были начаты раньше, чем по Y-хромосоме.
“Митохондриальная Ева”
Первые получившие известность исследования мтДНК представителей разных рас выявили общность происхождения всех ныне живущих людей по женской линии. В 1987 г. А.Уилсон и его коллеги из Калифорнийского университета в Беркли получили мтДНК от 147 представителей различных рас [1]. В исследуемом фрагменте они определили количество индивидуальных мутаций, а также их координаты и тип. Анализ показал, что все мтДНК возникли от одной предковой последовательности. По степени их разнообразия Уилсон оценил время, прошедшее с тех пор, как существовала предковая последовательность. Оно вычисляется как время схождения всех линий мтДНК в одну точку, называемую точкой коалесценции. В соответствии с его выводами, общая “праматерь”, к которой восходят все типы мтДНК современных людей, жила в Восточной Африке менее 200 тыс. лет назад. Установить последовательность возникновения мутаций в поколениях предков у обследованных индивидов тогда не удалось, но работы последних лет подтвердили выводы Уилсона.
Вопрос о прародине человечества дискутируется не одно десятилетие. Часть специалистов полагала, что человек возник в одном из регионов мира (наиболее часто упоминалась Африка) и затем расселился по всей Земле. Другая придерживалась так называемой мультирегиональной гипотезы, согласно которой предковый вид Homo erectus
превратился в Homo sapiens
в различных точках земного шара независимо. С появлением молекулярных данных значительный перевес получила “африканская” гипотеза.
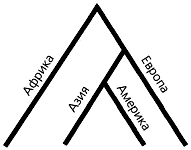 |
Филогенетическое древо популяций |
После работ Уилсона обладательницу предковой мтДНК окрестили “митохондриальной Евой”. Это сразу породило неверные толкования — будто все человечество произошло от одной женщины. На самом деле линии мтДНК современниц “Евы”, несомненно существовавших, до нашего времени не дошли. И произошло это не потому, что “Ева” была особенной. Концепция молекулярных часов построена на предположении о нейтральности большинства мутаций, т.е. подразумевает, что ничем особенным от соплеменниц “Ева” не отличалась. Просто генетическое разноообразие в популяции всегда уменьшается, если в череде поколений случаются резкие падения численности (прохождение “бутылочного горлышка”). А такое в истории человечества случалось не раз. Так что со временем все другие линии, кроме одной, исчезли по статистическим причинам.
Пока люди живут вместе, появляющиеся у них мутации распространяются по всей группе. Если же группы делятся, мутации в них накапливаются независимо. Число таких различий между группами пропорционально времени, прошедшему с момента их расхождения, что и позволяет датировать события популяционной истории. На основе изучения мтДНК населения разных регионов мира удалось выяснить пути и даты основных миграций прекрасной половины человечества.
Африканские предки
К большой радости генетиков реконструкция популяционной истории человечества по Y-хромосоме показала, что “Адам” — предок современных мужчин — жил примерно там же, где и “Ева”. Хотя данные, полученные при анализе Y-хромосомы, менее точны, чем митохондриальные, они также указывают на африканское происхождение человека и единую предковую популяцию. Молекулярные датировки времени ее разделения на ветви, ведущие к современным популяциям, зависят от методов оценки и, по данным разных авторов, колеблются от 30—40 тыс. лет назад (что противоречит палеонтологическим данным) до 180 тыс. лет назад. Как наиболее вероятный называют период от 57 до 135 тыс. лет назад [2].
Гипотезу африканского происхождения человека подтверждает ряд независимых исследований. Особенно интересны работы по изучению народов Южной и Восточной Африки — бушменов и готтентотов. Их языки содержат щелкающие звуки, нигде больше не встречающиеся, и относятся к так называемой койсанской группе (комбинация слов “кой-койн” — самоназвание готтентотов и “сан” — название бушменов), обособленно стоящей в системе языков мира. Они значительно отличаются от остальных африканских народов, в том числе и от своих соседей банту, не только лингвистически, но и антропологически: у них более светлая кожа, узкий нос, для женщин бушменов характерно избыточное отложение жира на ягодицах (стеатопигия), а для мужчин — особое строение половых органов. Отличия проявляются и в мтДНК: у бушменов обнаружена мутация, которая появилась одной из первых после возникновения человека как вида [3]. В Y-хромосоме некоторых бушменов также присутствует древняя мутация, не обнаруженная ни у кого из людей в других частях света, но найденная у обезьян. Вероятно, она возникла еще до разделения предковых линий человека и шимпанзе (5—7 млн лет назад) и непрерывно поддерживалась в части предковой популяции человека и шимпанзе. Возможно, эта мутация сохранилась только у бушменов потому, что их многочисленные предки в определенный момент истории заселяли значительную часть африканского континента. Впоследствии они были оттеснены на юг племенами, говорящими на банту. На это указывают и результаты изучения костных останков людей, найденных в Африке, — они сходны по типу с современными представителями койсанской группы.
Интересно, что популяционные различия по Y-хромосоме оказались в несколько раз выше, чем по мтДНК. Это говорит о том, что перемешивание генетического материала по женской линии более интенсивно, т.е. уровень миграции женщин почти на порядок выше, чем у мужчин [4]. И хотя эти данные на первый взгляд могут показаться удивительными (путешествия всегда считались прерогативой мужчин), их можно объяснить характерной для большинства человеческих обществ патрилокальностью (уходом жены в дом мужа). Брачные миграции женщин оставили более заметный след на генетической карте человечества, чем дальние походы Чингизхана или Аттилы [5].
Исследования ДНК неандертальцев
В 1997 г. генетик С.Пэбо сумел прочесть фрагмент мтДНК, выделенной из останков неандертальца, найденного в 1856 г. в Фельдгоферовской пещере близ Дюссельдорфа в Германии [6]. Эта пещера находится в долине Неандерталь (Neander Tal). В 1863 г. английский антрополог и анатом У.Кинг предложил для находки название Homo neanderthalensis *. Летом 2000 г. появилось сообщение другой группы ученых об исследовании второго образца неандертальской мтДНК из останков скелета ребенка, найденного в пещере Мезмайская на Северном Кавказе [7]. Во втором случае останки точно датированы радиоуглеродным методом — им 29 тыс. лет. Это — представитель одной из последних живших на Земле групп неандертальцев.
* Интересно, что название долины, данное в честь композитора XVII в. Иоахима Ноймана (Neumann — новый человек — по-гречески Neander), означает «новый человек».
Неандертальцы — группа гоминид, предки которых вышли из Африки раньше, чем предки современных людей. Неандертальцы населяли Европу и Западную Азию в период от 300 тыс. до 28 тыс. лет назад. Какое-то время они сосуществовали с человеком современного анатомического типа, расселившимся в Европе около 40 тыс. лет назад. Ранее на основе морфологического сравнения неандертальцев с человеком современного типа было предложено три гипотезы: неандертальцы — прямые предки человека; они внесли некоторый генетический вклад в генофонд Homo sapiens;
они представляли независимую ветвь, которую полностью вытеснил человек современного типа.
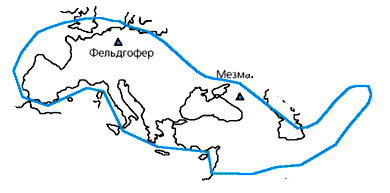 |
Места находок останков неандертальцев. Отмечено положение пещер Фельдгофер и Мезмай |
Результаты молекулярно-генетических исследований свидетельствуют в пользу третьей гипотезы. Обе неандертальские мтДНК имеют общие отличия от мтДНК людей, выходящие за границы внутривидового разнообразия H.sapiens. Это говорит о том, что неандертальцы — генетически отдельная, хотя и близкородственная человеку ветвь. По числу различий между их мтДНК время существования последнего общего предка человека и неандертальца оценивается в 500 тыс. лет. Палеонтологи считают, что предки неандертальцев появились в Европе около 300 тыс. лет назад, т.е. генетические линии, ведущие к человеку и неандертальцу, разделились раньше этой даты, как показывают датировки по мтДНК.
Таким образом, неандерталец эволюционировал в Европе одновременно с предком современного человека в Африке. Известно, что в Европе 40—50 тыс. лет назад расселился человек современного типа, а неандертальцы вымерли 28 тыс. лет назад. Неизвестно, какова связь этих событий — проиграл ли неандерталец в конкуренции с человеком или вымер по другим причинам.
Волны миграций и заселение континентов
Считается, что циклические изменения климата, происходившие с интервалом в десятки тысяч лет, играли существенную роль в эволюции и распространении всех видов, в том числе и человека. В периоды похолодания нарастала масса континентальных ледников, обитаемые природно-климатические зоны сдвигались на юг, уровень моря и озер снижался на сотни метров, увеличивалась площадь пустынь, съеживались тропические леса. С наступлением ледника в Европе сокращались обитаемые зоны и численность животных, сплошное “море” жизни распадалось на отдельные “озера”.
В периоды потепления повышались численность и разнообразие живых форм, виды расселялись на пригодные для жизни территории — из Африки в Азию и Европу через Суэцкий перешеек и в Европе — на освободившиеся из-под ледника территории. Таким образом, контакты африканской и евразийской биоты определялись глобальными циклами потепления—похолодания. Эти колебания приводили к значительным изменениям флоры и фауны всего Земного шара.
.
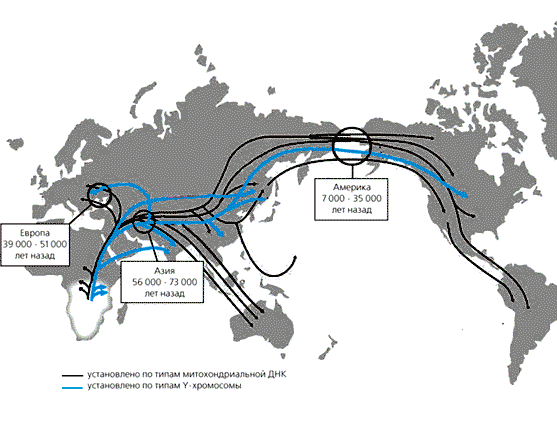
Пути и время расселения людей, установленные при анализе основных
материнских и отцовских (показано цветом) генетических линий
Исследования генетического разнообразия современных людей показали, что оно значительно ниже, чем, например, у шимпанзе, наших ближайших родственников в мире животных. Это означает, что человек как вид значительно позже шимпанзе прошел через “бутылочное горлышко” — резкое падение численности популяции, и/или это падение для человека было гораздо существеннее, чем для шимпанзе.
Различные группы генетиков пришли к выводу, что на протяжении последнего миллиона лет численность популяции прямых предков человека колебалась от 40 до 100 тыс. В период прохождения “бутылочного горлышка”, около 130 тыс. лет назад, значительно снизилось генетическо разнообразие предков человека, так как их общая численность сократилась до 10 тыс. индивидов и носители многих генетических вариантов исчезли [10]. Для сравнения — численность ныне живущих шимпанзе (а этот вид находится под угрозой исчезновения) составляет 100—200 тыс. особей.
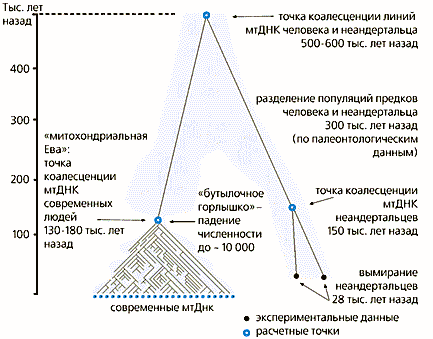
Схематическое представление популяционной истории человека и неандертальца на основе анализа мтДНК и палеонтологических данных. Линии мтДНК современных людей сходятся в точке (точка коалесценции), соответствующей снижению численности популяции (“бутылочному горлышку”) и отделяющей человека современного анатомического типа от архаических африканских гоминид. Разделение генетических линий (точка коалесценции) предшествует разделению популяций. Цветом показаны предполагаемые колебания численности популяций, установленные по уровню разнообразия ДНК современных людей.
Сравнительный анализ мтДНК разных популяций современных людей позволил предположить, что еще до выхода из Африки, около 60—70 тыс. лет назад (когда также наблюдалось снижение численности, хотя и не столь значительное), предковая популяция разделилась по крайней мере на три группы, давшие начало африканской, монголоидной и европеоидной расам [8]. В отечественной литературе их обозначают как экваториальная, азиатско-американская и евразийская расы, что более точно отражает их современное географическое распространение.
Часть расовых признаков, возможно, возникла позже как адаптация к условиям обитания. Это относится по крайней мере к цвету кожи — одному из наиболее значимых для большинства людей расовых признаков. Степень пигментации у человека генетически задана и, вероятно, в каждой популяции соответствует географической широте обитания. Пигментация обеспечивает защиту от солнечного облучения, но не должна препятствовать образованию, например, некоторых витаминов, предотвращающих рахит и необходимых для нормальной плодовитости [12].
На основе генетических данных постепенно проясняется картина заселения Азии, Европы и Америки. В недавно опубликованных работах определены частоты древних типов мтДНК и Y-хромосом, принесенных в Европу 40—50 тыс. лет назад первыми поселенцами, а также других, распространившихся позже, в том числе и тех, которые отражают экспансию земледельческих племен с Ближнего Востока 9 тыс. лет назад. И здесь генетические исследования пролили свет на еще один вопрос, также вызывавший на протяжении многих лет горячие споры.
Как распространяется культура? Перенимают ли люди традиции, технологии и идеи от представителей другой культуры (концепция культурной диффузии); или идеи и технологии путешествуют по миру только вместе со своими носителями, и смена культуры происходит одновременно со сменой населения (концепция демической диффузии)?
До недавнего времени преобладала концепция демической диффузии. Считалось, что земледельцы, пришедшие в Европу из Малой Азии около 10 тыс. лет назад, внесли основную долю в генофонд современных европейцев, вытеснив проживавшие в Европе палеолитические популяции [9]. Однако опубликованные в прошлом году, но уже общепризнанные работы [10, 11] показали, что генетическая доля мигрантов-земледельцев составляет в современном населении Европы не более 10—20%. Это значит, что относительно небольшое число земледельцев привнесло в палеолитическую Европу технические новации, и в результате на всей европейской территории сменился тип хозяйства и культуры.
Генетическое разнообразие народов России
В России изучение генетического разнообразия видов имеет очень давнюю традицию, восходящую к работам 20-х годов прошлого столетия основателей отечественной генетики А.С.Серебровского и С.С.Четверикова. Крупнейший итог популяционно-генетических иследований человека 80—90-х годов — пятитомное издание “Генофонд и геногеография народонаселения” под общей редакцией профессора Ю.Г.Рычкова из Института общей генетики РАН (Рычков и др., 2000). Его труды, выполненные в рамках фундаментальных научных программ РАН, РФФИ, а также программы “Приоритетные направления генетики”, и сейчас лежат в основе принципов подбора материала для современных исследований и формирования исходных гипотез. Ряд лабораторий, участников российской программы “Геном человека” (в том числе и наша лаборатория), изучает генетическое разнообразие народов европейской части России [12].
Линии мтДНК, попавшие в Европу в эпоху палеолита около 40—50 тыс. лет назад, составляют сейчас значительную часть мтДНК людей, населяющих территории от северо-запада Европы до Уральских гор. Наиболее четкие различия, обнаруженные между отдельными линиями мтДНК, указывают на их монголоидное или европеоидное происхождение. Мы обследовали русских и большинство народов Волжского бассейна и Урала. Выяснилось, что у всех народов, кроме башкир, по материнской линии преобладает европеоидный компонент. Градиент его падения проходит примерно с запада на восток и не коррелирует с языком этих народов. У русских (население Рязанской, Курской и Калужской областей), говорящих на языке индоевропейской группы, 97—98% материнских линий — европеоидные, а доля монголоидного компонента составляет 2—3%. Европеоидные линии преобладают также у народов финно-угорской языковой группы (мари, удмурты, коми) — от 80 до 94%, и у тюркоговорящих народов, проживающих в этом регионе. Так, 85% татар по материнской линии европеоидные, а чувашей 90%. Лишь у их восточных соседей, башкир, монголоидные линии составляют 65%. Значит, современная граница расселения европеоидов и монголоидов географически проходит примерно по Уралу, а популяционно-генетически — между башкирами, обитающими по обе стороны Уральского хребта, и их западными соседями - татарами. Повторим, что доля европеоидных и монголоидных линий не коррелирует с языком, на котором говорят эти народы.
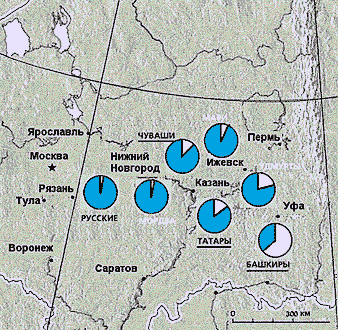
Материнские генетические линии у народов Волго-Уральского региона. Доля европеоидных линий мтДНК (цветной сектор) падает при продвижении с запада на восток, а доля монголоидных (белый сектор) возрастает. Исследованные этнические группы относятся к различным языковым семьям: индоевропейской (из славянских языков, относящихся к этой семье, приведены русские), уральской (финно-угорские языки; указаны белым шрифтом) и алтайской (тюркские языки; подчеркнуты)
Наши исследования показывают и разнообразные детали формирования отдельных народов. Например, монголоидные линии мтДНК у народов Волго-Уральского региона имеют разное происхождение: одни из них появились, вероятно, из Сибири, а другие — из Центральной Азии. Сочетание обнаруженных нами генетических линий образует мозаику, которая характеризует каждый из этих народов.
Проектов по изучению генетического разнообразия людей становится все больше. Они дают сведения, важные для здравоохранения, а также для реконструкции исторических событий. Сейчас известно, что многие мутации не нейтральны, скорость их накопления может быть различной для разных участков ДНК и на разных этапах эволюции. Поэтому абсолютные датировки, полученные на основе молекулярных методов, могут достаточно сильно различаться в зависимости от используемой системы анализа, и должны уточняться по мере развития экспериментальных методов и теоретических инструментов исследования.
В заключение можно предположить, что сложившиеся к настоящему времени представления о последовательности эволюционных и миграционных событий в истории человека как вида вряд ли сильно изменятся. Это, однако, не исключает сюрпризов при выявлении деталей формирования и взаимодействия разных популяций, приводивших к возникновению и смене языков и культур. Такие исследования помогут не только лучше понять причины, определившие современную структуру народонаселения Земли, но и предсказать тенденции этих процессов, что может быть крайне важным для выработки стабильных и сбалансированных отношений между народами в будущем.
Литература
1. Cann R.L., Stoneking M., Wilson A.C. // Nature. 1987. V.325. №6099. P.31—36.
2. Zhivotovsky L.A. // Mol. Biol. Evol. 2001 (in press).
3. Viglant L., Stoneking M., Harpending H. et al. // Science. 1991. V.253. P.1303—1307.
4. Seielstad M.T., Minch E., Cavalli-Sforza L.L. // Nat. Genet. 1998. V.20. P.278—280.
5. Stoneking M. // Nat. Genet. 1998. V.20. P.219—220.
6. Krings M., Stone A., Schmitz R.W. et al. // Cell. 1997. V. 90. №1. P.19—30.
7. Ovchinnikov I.V., Gotherstrom A., Romanova G.P. et al. // Nature. 2000. V.404. №6777. P.490—493.
8. Lahr M.M., Foley R.A. // Yearbook of physical anthropology. 1998. V.41. P.137—176.
9. Cavalli-Sforza L.L. Genes, peoples, and languages. N.Y., 2000.
10. Richards M., Macaulay V., Hickey E. et al. // Am. J. Hum. Genet. 2000. V.67. P.1251—1276.
11. Underhill P., Cavalli-Sforza L.L., Oefner P.J. // Nat. Genet. 2000. V.26. P.358—361.
12. Orekhov V., Poltoraus A., Zhivotovsky L.A. et al. // FEBS Lett. 1999. V.445. №1. P.197—201.