| Скачать .docx | Скачать .pdf |
Доклад: Доклад: Аварийная сигнализация в живом организме
Человек живет в постоянном контакте с внешней средой, получая информацию об окружающем мире с помощью специализированных сенсорных систем, воспринимающих механические, термические, акустические, электромагнитные (световые) и химические сигналы. Благодаря работе этих систем мы можем, например, любоваться светом звезд, наслаждаться пением птиц, ароматом цветов и т.д. Однако всем наверняка приходилось испытывать и другое, вовсе неприятное, ощущение — боль, возникающую в результате какого-либо вредного для организма воздействия. Ответственна за это так называемая ноцицептивная система (от лат. noсeo — вредить). И хотя боль дает мало сведений об окружающем нас мире, тем не менее она столь же необходима человеку для нормальной жизни, как осязание или обоняние, так как предостерегает о внешних или внутренних опасностях, грозящих нашему телу. Несмотря на то, что боль во многом сравнима с другими чувствами, она все же обладает характерными особенностями. Чтобы помочь страдающему от боли человеку, надо хорошо понимать ее специфические черты.
В отличие от избирательных сенсорных систем для ноцицепции не существует специализированного болевого стимула (боль возникает при ушибе, ожоге, укусе пчелы, и т. д.). Нет и особого, анатомически выделенного органа, подобного, например, органу слуха или зрения. Наконец, болевое ощущение может быть вызвано на любом участке тела, соответственно в организме нет каких-либо специфических болевых нервов, подобных зрительному или обонятельному. Это — своеобразная аварийная сигнализация живого организма, которая включается в критической ситуации. И организм (еще до того как мы осознаем случившееся и примем решение) немедленно реагирует — срабатывает защитный рефлекс, выражающийся в многокомпонентной оборонительной реакции. Достаточно вспомнить, как человек отдергивает обожженную руку или меняет позу при боли в позвоночнике (моторный компонент), что нередко сопровождается криком или агрессией (поведенческий компонент). При этом обычно изменяются кровяное давление и ритм дыхания, учащается пульс, расширяются зрачки (реакции вегетативной нервной системы) и т.д.
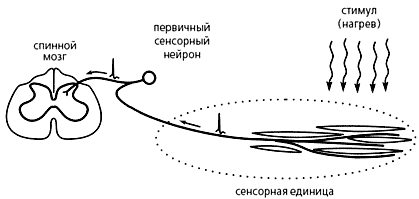 |
Путь болевого сигнала
в живом организме. |
Таким образом, живой организм может распознать внешние воздействия (стимулы), способные причинить ему вред. Порог обнаружения таких воздействий весьма близок к той силе стимула, при которой действительно происходит повреждение ткани, на что впервые обратил внимание знаменитый английский физиолог Ч.С.Шеррингтон [1]. Казалось бы, любые вредящие стимулы должны вызывать защитный рефлекс, но, к сожалению, наш организм еще недостаточно совершенен. Существуют вредные воздействия, которые не выявляются нервной системой (например, радиация), а некоторые безвредные стимулы (электрический ток или ультразвук умеренной интенсивности), напротив, вызывают боль. Учитывая это, в физиологии принято называть ноцицептивными только те стимулы, которые вызывают защитный рефлекс. Защитный рефлекс, ноцицептивная система и вредящие стимулы — ключевые понятия теории боли.
Ноцицептивная система оказалась существенно сложнее для изучения, чем избирательные сенсорные системы. Отсутствие специфического болевого стимула, специальных болевых нервов и специализированного органа болевой чувствительности объясняет значительное отставание в исследовании механизмов работы этой системы по сравнению с другими. Раскрытие основных принципов и структурных элементов ноцицептивной системы — итог более чем столетних исследований. Оказалось, что в восприятии и обработке информации о вредящих воздействиях участвуют различные отделы нервной системы — от первичных сенсорных нейронов до определенных структур головного мозга. Мы рассмотрим только начальное, периферическое звено ноцицептивной системы, но сначала еще несколько слов о терминах и основных понятиях.
Ноцицептивные сигналы возникают в окончаниях первичных сенсорных нейронов и поступают в спинной мозг сначала по периферической, а затем центральной ветвям их аксонов. Теоретически биполярная структура первичного сенсорного нейрона допускает преобразование импульсных последовательностей на этом пути. Учитывая это, физиологи ввели понятие “сенсорная единица” (специфический рецептор в избирательной сенсорной системе и ноцицептор — в ноцицептивной), которое объединяет периферическую ветвь аксона первичного сенсорного нейрона (афферентное волокно), ее тканевое окончание (терминаль) и возможные претерминальные клетки-сателлиты, функционально связанные с терминалью. Сенсорную единицу можно представить как некий функциональный модуль, для которого входной сигнал — внешний стимул, а выходной — последовательность нервных импульсов в периферической ветви аксона первичного сенсорного нейрона.
Периферическая ноциология начала развиваться в XIX в., когда уровень техники был еще недостаточен для решения ее задач. В 1838 г. немецкий физиолог И.П.Мюллер предположил, что каждому ощущению, включая боль, соответствуют определенные специфические нервные волокна (в современных терминах — первичные сенсорные нейроны и ноцицепторы). В конце XIX в. гистолог М.Бликс обнаружил специализированные точки кожи для определенных видов стимулов, физиолог М.Фрей установил различную чувствительность малых участков кожи к точечным механическим стимулам [2, 3]. Эти работы привели к представлению о сенсорной гетерогенности кожи и послужили основой для формулирования теории специфичности боли. Согласно этой теории, существуют специализированные кожные нервные волокна, чувствительные окончания (ноцицепторы) которых возбуждаются только вредящими стимулами.
В XIX в. появилась и другая, альтернативная, теория боли — теория интенсивности, согласно которой защитный рефлекс и боль возникают при чрезмерном раздражении любого нерва. Сформулировал ее в 1895 г. немецкий врач А.Гольдшайдер, заметивший, что при некоторых заболеваниях происходит пространственное и временнOе суммирование ноцицептивных стимулов [4]. Теория интенсивности предполагает, что все сенсорные единицы выполняют двойную роль как специфических рецепторов (например, тепловых), так и ноцицепторов.
В XX в. обе конкурирующие теории были экспериментально проверены с помощью развитых к тому времени гистологических методов и новых электронных приборов. Английские гистологи школы Г.Уэдделла не выявили связи между структурой и функцией нервных окончаний кожи человека [5]. Оказалось, что все разнообразие кожных ощущений передается нервными окончаниями всего двух типов — инкапсулированными и свободными. Диссонанс между гаммой кожных ощущений и скудным набором структурных типов сенсорных терминалей особо заметен в волосистой коже, где инкапсулированных терминалей почти нет. Гистологи заметили и другой важный факт: большинство микроскопических участков кожи снабжено свободными нервными окончаниями различных аксонов. Это означает, что даже строго локализованный стимул одновременно возбуждает различные сенсорные единицы.
Результаты гистологических исследований послужили основанием как для критики теории специфичности, так и для развития теории интенсивности, что привело к созданию теории паттерна (нейроимпульсного узора) Дж.Нейфа [6]. Согласно этой теории, ноцицептивному (как и любому другому) стимулу соответствуют определенные импульсные последовательности (паттерны) в группах нервных волокон. Как и теория интенсивности, теория паттерна не предполагает существования специализированных ноцицепторов для распознавания повреждающих стимулов. Считается, что одно и то же волокно выступает в разных ипостасях, внося вклад в суммарный импульсный узор, приводящий к тому или иному ощущению.
К сожалению, структурную однородность нервных окончаний гистологи без каких-либо оснований распространили и на их функциональную однотипность, поэтому критика теории специфичности оказалась некорректной. В действительности же внешне одинаковые нервные окончания могут резко отличаться по реакции на различные воздействия. Однако выяснилось это, лишь когда удалось зарегистрировать сигналы в нервных волокнах.
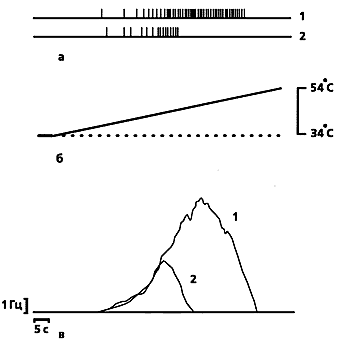
| Типичная запись разрядов кожного ноцицептора (а) кошки при линейно-нарастающем нагреве (б), полученная в наших опытах [15]. Кривая (в) показывает среднюю частоту разрядов и отражает характерные особенности ноцицептивных сигналов индивидуальных С-ноцицепторов, которые представляют собой ограниченную последовательность из нескольких десятков нервных импульсов, следующих с частотой, не превышающей 20 Гц. | 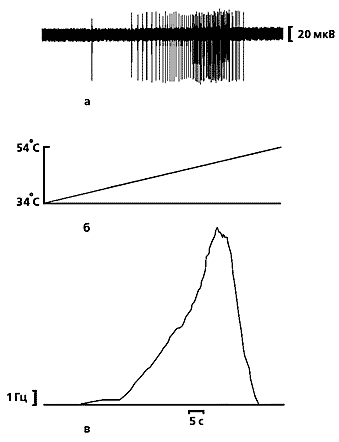 |
Изобретение осциллографа совершило революцию в нейрофизиологии. В 1921 г. с его помощью американские исследователи, физиолог Г.Гассер и физик Г.Ньюкомер, впервые “увидели” нервные импульсы [7]. Оказалось, что нерв состоит из волокон, значительно (более чем в 100 раз) отличающихся по скорости проведения нервных импульсов. Наибольшую скорость проведения имели толстые миелиновые Аb -нервные волокна диаметром до 20 мкм, а самую малую — тонкие миелиновые Аd -волокна и безмиелиновые (безмякотные) С-волокна диаметром около 1 мкм. Применение осциллографа позволило зарегистрировать суммарные сигналы отдельных групп нервных волокон и выявить роль этих групп в передаче ноцицептивных сообщений.
В 1926 г. англичанин Э.Эдриан и швед Ю.Цоттерман впервые получили записи сигналов в одиночных толстых миелиновых афферентах скелетной мышцы кошки [8]. Был установлен принципиальный факт: “чрезмерное раздражение” Аb -афферентов кожи, вызывающее разряд импульсов с максимальной для этих волокон частотой, не стимулирует возникновение защитного рефлекса. Так на примере толстых миелиновых волокон была доказана несостоятельность теории интенсивности. Что касается тонких С-волокон, то, как оказалось, они играют особую роль в передаче ноцицептивных сигналов. Эдриан и Цоттерман выяснили, что импульсы в группе С-афферентов кожи возбуждаются при травмирующих механических, химических и температурных воздействиях [9, 10]. Д.Кларк, Дж.Хьюз и Г.Гассер установили, что при силе электрического стимула, достаточной для возбуждения С-афферентов, возникал защитный рефлекс, причем он развивался и при блокаде толстых миелиновых Аb -афферентов [11].
Таким образом, в первой половине XX в. физиологи выяснили, что ноцицептивные сигналы передаются по наиболее тонким афферентным волокнам группы С. Несколько позже оказалось, что этот вывод справедлив и для самых тонких из миелиновых волокон (группы Аd ). Означало ли это, что тонкие афференты (волокна групп С и Аd ) те же специализированные проводники ноцицептивных сигналов, существование которых предполагает теория специфичности? В 1957 г. У.Дуглас и Дж.Ритчи разработали метод встречных импульсов, с помощью которого обнаружили выраженное возбуждение группы С-афферентов не только ноцицептивными, но и слабыми механическими стимулами [12]. В связи с этим представление о том, что тонкие афференты передают информацию только о ноцицептивных стимулах, оказалось ошибочным. В результате противоборство двух классических теорий сохранилось, однако область их противостояния сузилась и стала относиться теперь только к тонким афферентам.
Следующий шаг в споре двух теорий мог быть сделан только на основе опытов с регистрацией сигналов индивидуальных С-афферентов. Сложность подобных опытов связана с исключительно малым диаметром этих нервных волокон (около 1 мкм) и их слабыми электрическими сигналами. Методические проблемы были решены в 30—50-х годах благодаря развитию микрохирургии нервов, а также совершенствованию техники регистрации электрических сигналов от микропучков нервных волокон. Значительный вклад в решение этих проблем внесли европейские (Эдриан и Цоттерман), японские (Дж.Като и И.Тасаки) и индийские (школа А.Пайнтала) нейрофизиологи. В 1955 г. британский физиолог А.Игго получил устойчивые записи сигналов индивидуальных С-афферентов, применив для изоляции микропучков нервов жидкий диэлектрик (вазелиновое масло) вместо воздуха [13].
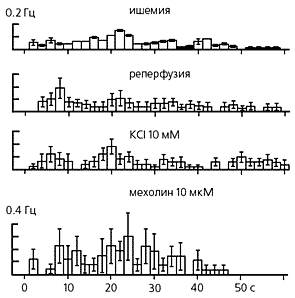 |
Усредненные гистограммы ответов кожных С-ноцицепторов, вызванных неноцицептивными стимулами: остановкой кровотока в подкожной артерии (ишемия), возобновлением кровотока (реперфузия), а также внутриартериальным введением калия (в малой концентрации) и мехолина. |
В 1969 г. американские физиологи П.Бессу и Э.Перл зарегистрировали сигналы таких индивидуальных кожных С-афферентов кошки, чувствительные окончания которых возбуждались различными, но исключительно ноцицептивными (механическими, термическими и химическими) стимулами [14]. Полимодальность этих сенсоров и их избирательная чувствительность к ноцицептивным стимулам идеально соответствовали предполагаемым свойствам ноцицепторов. Открытие ноцицепторов подвело итог длительному этапу в развитии ноциологии, и теория специфичности одержала убедительную победу.
Выяснение характеристик ноцицептивных сигналов имеет исключительно важное значение для медицины, поскольку именно эти сигналы должны подавляться при обезболивании. Казалось бы, нейрограммы помогают ответить на этот вопрос, однако различные виды вредящих стимулов вызывают в ноцицептивных афферентах разные паттерны разрядов. Так, разряды в ноцицепторах, вызываемые давлением на кожу, продолжаются всего 1—3 с, в то время как тепловые или химические стимулы (например, ацетилхолин или гистамин) вызывают разряды, длящиеся около минуты.
Означает ли такое разнообразие ответов, что любой разряд в ноцицептивных афферентах — болевой сигнал? Если так, местное обезболивание может быть достигнуто только при полной блокаде сигналов практически во всем нервном стволе, поскольку существующие местные анестетики “не отличают” С-волокна ноцицепторов от волокон других сенсорных единиц. Такой вид местного обезболивания действительно применяется при хирургических операциях. Однако полная блокада требует весьма высоких концентраций анестетика, что недопустимо, например, при длительном обезболивании. Можно ли осуществить местное обезболивание без тотального блокирования нервных проводников? Если допустить, что любой разряд импульсов в ноцицептивных афферентах — болевой сигнал, тогда для обезболивания нужны были бы анестетики, способные избирательно и полностью блокировать ноцицепторы, что пока практически невыполнимо. Напротив, если в ноцицептивных афферентах возможна импульсная активность, не ведущая к боли, то обезболивания можно было бы достичь частичным угнетением ноцицептивных нейронов (соответственно применяя анестетик в меньших концентрациях). При этом проводимость сигналов в других сенсорных единицах была бы также частично угнетена — однако не подавлена полностью, что очень важно. Дело в том, что во многих случаях сохранение связи пораженного органа с центральной нервной системой имеет принципиальное значение для лечения.
Сравнительно недавно нам удалось, пока еще в эксперименте, решить эту проблему. Принципиальную возможность возбуждения ноцицепторов заведомо неноцицептивными воздействиями нельзя выяснить с помощью тепловых или механических стимулов, поскольку они возбуждают ноцицепторы лишь при такой силе, когда их трудно отличить от ноцицептивных. Исходя из этого, мы выбрали химические воздействия, о которых было заведомо известно, что они неноцицептивные: например, внутриартериальное введение калия в малой концентрации и мехолина —агониста M-холинорецепторов. Согласно теории специфичности, подобные вещества не должны возбуждать ноцицепторы. Однако в результате эксперимента выяснилось, что не только эти, но и некоторые другие неболевые раздражители, например остановка кровотока в подкожной артерии (ишемия) и его возобновление (реперфузия), способны возбудить кожные С-ноцицепторы кошки. Любопытно, что ноцицептивные химические стимулы (например, высокие концентрации ацетилхолина — агониста N-холинорецепторов) вызывали в тех же сенсорных единицах более интенсивные разряды, чем неболевые агенты (например, мехолин в любых концентрациях).
| Ответы кожных С-ноцицепторов, вызванные болевым (в данном случае: “болевыми” концентрациями ацетилхолина, слева) и неболевым (мехолином даже при увеличении его концентрации до 500 мкМ) химическими стимулами. Ноцицептивные стимулы всегда вызывают высокочастотные разряды (выше 2 Гц). | 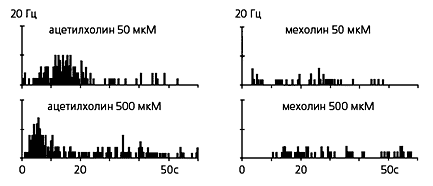 |
Таким образом, стала понятна природа ноцицептивного сигнала: им оказалось высокочастотное возбуждение (вплоть до 20 Гц) ноцицепторов. Примерная частотная граница, отделяющая ноцицептивные разряды от субноцицептивных, составляет 2 Гц.
Итак, теория специфичности одержала лишь промежуточную победу, и исторический спор двух теорий приостановлен лишь на время. Если будет установлена физиологическая роль низкочастотных (неболевых) разрядов ноцицепторов и, возможно, определено ощущение, вызываемое этими разрядами, то в этом случае придется вернуться к теории интенсивности, теперь уже только для многофункциональных сенсорных единиц. В качестве кандидатов на роль специфических (неболевых) стимулов для полимодальных афферентов могут выступать тканевые метаболиты (в соответствии с теорией тканевых интероцепторов знаменитого российского физиолога В.Н.Черниговского [16]). Ощущения, вызываемые субноцицептивными разрядами тканевых интероцепторов, могли оказаться зудом или мышечной усталостью. “Чрезмерное” (высокочастотное) возбуждение тканевых интероцепторов являлось бы ноцицептивным сигналом. Подобное развитие ноциологии стало бы триумфом теории интенсивности. В то же время возможен и менее увлекательный сценарий дальнейшего развития теории боли: субноцицептивные разряды ноцицепторов могут оказаться всего лишь низкочастотным информационным шумом, не играющим никакой сенсорной роли. В этом случае вновь “победит” теория специфичности.
Для практической медицины наши выводы не менее важны. Существование низкочастотных (неболевых) разрядов в ноцицепторах позволяет добиться идеального местного обезболивания без полного угнетения импульсной активности в нерве [17]. И вот как можно реализовать эту возможность. В наших опытах лекарственный препарат N-пропилаймалин, введенный подкожно, “срезал” высокочастотную составляющую в ответе кожного С-ноцицептора на болевой нагрев. Эта своеобразная низкочастотная фильтрация сигналов ноцицепторов может быть прообразом такого способа местного обезболивания, при котором подавляется боль, но сохраняется связь пораженного органа с центральной нервной системой, что исключительно важно для лечения.
 |
Избирательное блокирование N-пропилаймалином (0.01%) высокочастотных разрядов кожного С-ноцицептора при тепловом раздражении. Вверху (а) показаны гистограммы разрядов до (1) и после (2) подкожного введения препарата. Ниже — температура рецепторного поля (б) и соответствующие кривые средней частоты разрядов (в). |
Мы рассказали лишь о некоторых проблемах современной ноциологии. Только новые достижения в технике помогут, как и сто лет назад, ответить на очередные вопросы, поставленные открытием ноцицепторов и обнаружением их возбудимости неболевыми стимулами. Главные успехи ноциологии впереди: они будут основаны на возможности одновременной регистрации индивидуальной импульсной активности практически всех волокон достаточно крупного нерва у бодрствующего человека. Сегодня это лишь фантазия — но не бOльшая, чем рассуждения о природе “животного электричества” во времена Луиджи Гальвани.
Литература
1. Шеррингтон Ч. Интегративная деятельность нервной системы. Л., 1969.
2. Blix M. // Z. Biol. 1884. Bd.20. S.141—156.
3. Frey M. //Berichte Verhandl. Gesellsch. Wissensch. 1894. Bd.46. S.185—196.
4. Гольдшайдер А. О боли с физиологической и клинической точки зрения. М., 1895.
5. Weddell G. // Ann. Rev. Psychol. 1955. V.6. P.119—136.
6. Nafe J.P. // Am. J. Psychol. 1927. V.39. P.367—389.
7. Gasser H.S., Newcomer H.S. // Am. J. Physiol. 1921. V.57. P.1—26.
8. Adrian E.D., Zotterman Y. // J. Physiol. 1926. V.61. P.151—171.
9. Adrian E.D. // Proc. Roy. Soc. Lond. 1931. V.109. P.1—18.
10. Zotterman Y. // J. Physiol. 1939. V.95. P.1—28.
11. Clark D., Hughes J., Gasser H.S. // Am. J. Physiol. 1935. V.114. P.69—76.
12. Douglas W.W., Ritchie J.M. // J. Physiol. 1957. V.139. P.385—399.
13. Iggo A. // J. Physiol. 1955. V.128. P.593—607.
14. Bessou P., Perl E.R. // J. Neurophysiol. 1969. V.32. P.1025—1043.
15. Ревенко С.В., Байдакова Л.В., Ермишкин В.В., Мангушева Н.А., Селектор Л.Я. // Нейрофизиология. 1992. Т.24. №5. С.517—529.
16. Черниговский В.Н. Интероцепция. Л., 1985.
17. Боровикова Л.В., Боровиков Д.В., Ермишкин В.В., Ревенко С.В.
// Сенс. системы. 1997. Т.11. №2. С.107—117.