| Скачать .docx | Скачать .pdf |
Доклад: Доклад: Органы кроветворения и иммуногенеза
ОРГАНЫ КРОВЕТВОРЕНИЯ И ИММУНОГЕНЕЗА
К органам кроветворения и иммуногенеза относят: красный костный мозг, тимус, лимфатические узлы, селезенку, миндалины, а также лимфоидные образования слизистой оболочки пищеварительной, половой, дыхательной и выделительной систем. Их подразделяют на центральные и периферические органы. Центральные органы включают в себя красный костный мозг и тимус (а также клоакальную сумку у птиц). В красном костном мозге образуются все форменные элементы крови и в том числе предшественники Т- и В-лимфоцитов. В тимусе, главном органе лимфоцитопоэза, размножаются различные клоны Т-лимфоцитов; в клоакальной сумке птиц — В-лимфоциты (антиген-независимое размножение).
К периферическим иммунным органам относят лимфатические и гемолимфатические узлы, селезенку и лимфоидные образования, локализующиеся в стенке пищеварительной трубки и в других органах. В них пролиферируют и дифференцируются Т- и В-лимфоциты, мигрирующие из центральных органов иммуногенеза. Под влиянием антигенов лимфоциты трансформируются в эффекторные и регуляторные клетки, способные выполнять иммунологическую защиту. Здесь же выбраковываются и утилизируются погибающие клетки крови.
Все органы кроветворения и иммуногенеза функционально тесно связаны друг с другом и действуют согласованно, обеспечивая постоянство клеточного состава крови и индивидуальный иммунологический статус организма. В регуляции кроветворения и иммуногенеза участвуют нервная и эндокринная системы. Не менее важна роль и микроокружения гемопоэтических клеток, например ретикулярной ткани, которая не только образует механический каркас, но и создает микросреду, стимулирующую пролиферацию, созревание и мигра![]()
цию гемопоэтических и лимфоидных клеток. Наличие в кроветворна органах специализированных фагоцитарных элементов, способна'* очищать протекающую через них кровь или лимфу от инородных эл'* ментов (бактерий, протистов, остатков погибших клеток), также сви" детельствует о защитной функции данных органов.
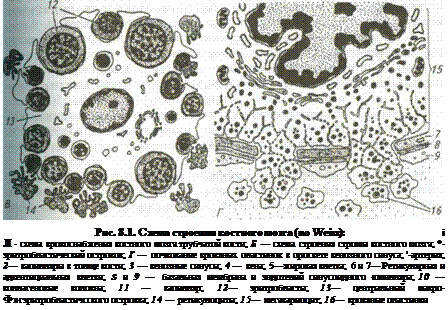 |
Костный мозг. У взрослых животных его подразделяют на красный и желтый, что обусловлено разной структурой содержимого мозга' в первом в большом количестве присутствуют эритроциты, во втором преобладают жировые клетки.
Красный костный мозг (medullaossiumrubra). Он располагается в губчатом веществе костей свода черепа, ребер, грудины, в телах позвонков в гребне подвздошной кости, а также в эпифизах трубчатых костей и пред.! ставлен миелоидной тканью, состоящей из трехмерной сети клеток ретикулярной ткани (textusreticularis), ретикулярных и коллагеновых волокон формирующих строму, а также клеток крови, находящихся на различных стадиях развития (рис. 8.1, см. также цв. вклейку).
Миелоидная ткань локализуется в костных полостях между костными перекладинами. Она образована сетью ретикулярных клеток, между которыми обнаруживают большое количество тонкостенных фенест-рированных синусоидных капилляров (vascapillaresinusoideum). Через стенки последних новообразованные клетки крови поступают в общий кровоток. В миелоидной ткани находится основная популяция стволовых кроветворных клеток (СКК), являющихся родоначальниками трех самостоятельных линий форменных элементов крови: эритроцитопо-этической, гранулоцитопоэтической и мегакариоцитарно-тромбоци-топоэтической. В ней проходят начальные стадии развития популяции Т- и В-лимфоцитов.
Среди гемопоэтических клеток красного костного мозга выделяют: стволовые и полустволовые клетки, морфологически сходные с малыми лимфоцитами и трудно идентифицируемые обычными гистологическими методами; властные формы (проэритробласты, промиело-циты, метамиелоциты, пролимфоциты, промоноциты, мегакари-областы, мегакариоциты); дифференцирующиеся и зрелые форменные элементы крови. Соотношение зрелых и незрелых форм в красном костном мозге и в периферической крови служит важным диагностическим показателем нормы или патологии. В норме в кровоток из красного костного мозга попадают только зрелые форменные элементы крови и некоторое количество клеток, мигрирующих в другие органы, например, в лимфоидные.
Гемопоэтические клетки, как правило, образуют островки, локализующиеся между синусоидными капиллярами. Эритробласты в процессе созревания формируют розетки вокруг макрофага, содержащего в цитоплазме железо, которое используется эритробластами для синтв' за гемоглобина. Другая функция макрофагов — поглощение выталкиваемых из нормоцитов ядер.
Клетки эритробластического ряда — проэритробласты (proerythro-blastus) характеризуются округлым или овальным ядром с мелким диффузным хроматином, крупным ядрышком и слегка базофильной цитоплазмой. Располагаются проэритробласты в очажке небольшими группами и после интенсивной пролиферации превращаются в более мелкие клетки с резко базофильной цитоплазмой — базофильные э Ритробласты (erythroblastusbasophilicus), тоже интенсивно размножающиеся. По мере синтеза гемоглобина цитоплазма клеток начинает воспринимать кислые красители — клетки превращаются в полих матофильные эритробласты (etythtroblastuspolichromathophilicus) в paxпоследних ядрышко отсутствует, но они продолжают активно xi*' лиферировать. После дифференцировки и накопления гемоглоби°" клетки переходят в стадию оксифильных эритробластов (erythrobla^ acidophilicus), теряют способность к размножению, ядра в них пикн тизируются и элиминируются из клетки. Клетка превращается вначал" в ретикулоцит, а затем в зрелый эритроцит. Продолжительность жизн эритроцитов в периферической крови, например, у свиньи составчЯе ! в среднем 70 дней, у крупного рогатого скота и овцы 50...60. В амоп/ ном веществе островков эритропоэза преобладают гликопротеиды ко личество которых уменьшается по мере созревания клеток и увеличе" ния их подвижности.
Между эритроидными островками встречаются дифференцирую.-щиеся клетки гранулоцитарного ряда (базофилы, эозинофилы, нейт-рофилы), а также группы жировых клеток. В островках миело- или гранулоцитопоэза в аморфном веществе преобладают протеогликаны количество которых уменьшается по мере созревания клеток. В процессе пролиферации и дифференцировки уменьшаются размеры миелоидных клеток, изменяется форма их ядер — от округлой до сегментированной, в цитоплазме накапливается специфическая зернистость. Общая продолжительность жизни нейтрофильных гранулоци-тов4...8 дней, эозинофилов 8... 12 дней, а лимфоцитов 10...15 дней.
Кровяные пластинки образуются в результате фрагментации цитоплазмы на отдельные участки особых гигантских полиплоидных клеток — мегакариоцитов, тесно контактирующих со стенкой синусоид-ных капилляров.
Наиболее трудно идентифицировать (наряду со стволовыми и полустволовыми предшественниками) костномозговые лимфоидные элементы (предшественники Т- и В-лимфоцитов и моноцитов), чаще встречающиеся вблизи капилляров. Наиболее интенсивное кроветворение отмечают вблизи эндоста, где концентрация стволовых клеток втрое выше, чем в центре костномозговой полости.
Желтый костный мозг (medullaossiumflava). Он находится в диафи-зах трубчатых костей и представлен преимущественно жировыми клетками, содержащими пигменты типа липохромов. Небольшое количество адипоцитов постоянно встречается и в красном костном мозге.
Тимус (зобная, или вилочковая, железа). В тимусе {thymus) происходит антигеннезависимая, генетически запрограммированная пролиферация и дифференцировка Т-лимфоцитов из Т-предшествеников (прекурсоров), мигрирующих из красного костного мозга. Тимус контролирует все иммунные реакции.
Орган появляется в эмбриогенезе достаточно рано. Источником его развития служит эпителий, прорастающий в виде трубок в подлее' щую мезенхиму из 3-го и 4-го жаберных карманов. Эпителиальна*
адка инфильтрируется лимфоцитами. Таким образом, в плане раз-за1СЛ я и тканевого строения, тимус представляет собой лимфоэпители-ВИТ нЫЙ орган, относящийся к группе бранхиогенных желез. Эпители-
ная основа органа проявляет черты эндокринной железы особенно ^ко в период внутриутробной жизни.
Тимус покрыт соединительнотканной капсулой, продолжающейся перегородки, содержащие сосуды и разделяющие доли железы на не-В лно отграниченные дольки. В зависимости от вида животного коли-П °ство долей в тимусе варьирует: у жвачных и свиньи железа состоит из Ч !ух долей — грудной непарной и шейной парной; у лошади и собаки щейная часть тимуса развита слабо или вовсе отсутствует, а развита только грудная непарная часть.
Долька состоит из трехмерной сети отросчатых эпителиоретикуляр-ньгх клеток, образующих строму органа. В петлях сети располагаются лимфоциты (тимоциты). В каждой дольке выделяют корковое и мозговое вещество. Их соотношение может меняться в зависимости от функционального состояния иммунной системы. В корковом веществе дольки сконцентрировано большое количество лимфоцитов, плотно прилегающих друг к другу, что придает ему более темную окраску на гистологических срезах. Мозговое вещество выглядит более светлым из-за того, что в нем существенно меньше лимфоцитов и на их фюне выделяются ретикулоэпителиальные клетки и тимусные тельца, или тельца Гассаля (рис. 8.2).
В корковом веществе выделяют субкапсулярную зону, где находятся крупные лимфоидные клетки — лимфобласты, или предшественники Т-лимфоцитов, мигрирующие сюда из красного костного мозга. Под влиянием гемопоэтических факторов, выделяемых эпителиальными ретикулоцитами, лимфобласты активно пролиферируют и превращаются в Т-лимфоциты, часть из которых переходит во внутреннюю зону коры, где они продолжают активно размножаться, дифференцируются, приобретая один из маркеров Т-клеток (Т-киллер, Т-хелпер, Т-суп-рессор), затем зрелые клетки перемещаются в мозговое вещество.
Эпителиоретикулярным клеткам свойственно овальное или округлое крупное светлое ядро с 2...3 ядрышками. В цитоплазме выявляют комплекс Гольджи, мелкие митохондрии, фрагменты ГлЭПС и секреторные вакуоли диаметром 0,5... 1,5 мкм. Своими отростками указанные клетки охватывают тимоциты, создавая микроокружение, необходимое для их деления и созревания. В корковом веществе различает: эпителиоретикулоциты следующих типов: секреторные клетки, вырабатывающие биологически активные вещества, необходимые для с °зревания тимоцитов; «клетки-няньки», включающие в свою цитоплазму до нескольких десятков тимоцитов, изолируя их от окружающей среды и, по-видимому, участвуя в их селекции; периваскулярные ^тки, охватывающие своими отростками сосуды и участвующие в Формировании гемато-тимического барьера.
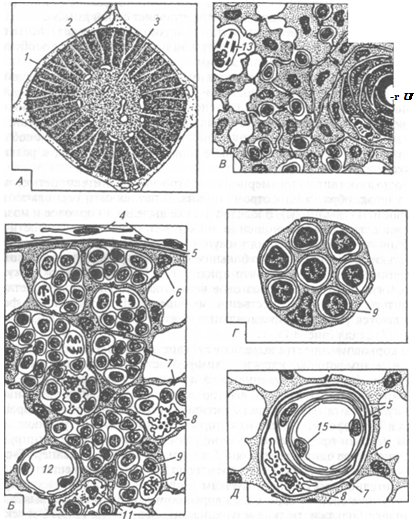 |
Рис. 8.2. Тимус (по Olah, Rohlich):
А — общий вид долек; Б — кора; Я — мозговое вещество; Г— клетка-нянька из наружной части коры; Д— строение гемато-тимического барьера; / — капсула; 2— кора; 3 — мозговое вещество; 4— фибробласт; 5— базальная мембрана; 6—эпителиальная строма; 7—десмосомы между эпителиальными клетками; 8— макрофаг; 9— лимфоцит; 10— лимфобласт в процессе митоза; // — интердигитирующая клетка; 12—просвет сосуда; 13—миоидная клетка; 14—центр ороговения тельца Гассаля; /5— клетка эндотелия
Однако не все лимфоциты, образовавшиеся в корковом веществе, выходят в циркуляторное русло, а лишь те, которые прошли «обучение» и, встретившись с чужеродными антигенами, приобрели специ-
ические мембранные рецепторы. Лимфоциты, несущие на своей поверхности рецепторы к собственным антигенам организма, погибают в тимусе. В противном случае такие лимфоциты, попав в кровоток, могут вызвать аутоиммунную реакцию (начнут атаку против собственных белков и клеток). Поэтому лимфоциты, образующиеся в коре тимуса, изолированы от антигенов прочным гематотканевым барьером, в состав которого входят: эндотелиоциты капилляров, их базальная мембрана, перикапиллярное пространство с соединительной тканью и активированными лимфоцитами и макрофагами, а также ретикуло-эпителиальные клетки стромы, окруженные базальной мембраной и связанные друг с другом десмосомами. Кроме того возможность попадания чужеродных антигенов предотвращается отсутствием в тимусе лимфатических сосудов. Барьер характеризуется избирательной способностью по отношению к антигену. При нарушении гематотканевого барьера в коре тимуса начинают обнаруживать единичные плазматические клетки, зернистые лейкоциты и тканевые базофилы.
Более светлый вид мозгового вещества дольки по сравнению с корковым обусловлен не только меньшей численностью и плотностью тимоцитов, но и почти полным отсутствием пролиферирующих лимфоцитов: их здесь в 10...15 раз меньше, чем в коре долек. В мозговом веществе большая концентрация ретикулоэпителиоцитов. Отличительная особенность тимуса — это наличие слоистых эпителиальных телец (corpusculumthymi), или телец Гассаля, производных ретикулоэпителиоцитов. На периферии телец находятся функционально нормальные уплощенные эпителиальные клетки, а в центре — дистрофически измененные. Количество и размеры слоистых телец увеличиваются с возрастом. В соединительной ткани, сопровождающей сосуды, а иногда и в паренхиме медуллы встречаются тучные клетки, а также клетки миелоидного ряда.
Помимо Т-лимфоцитов из тимуса в кровь поступают биологически активные гормоноподобные вещества, которые содействуют созреванию Т-лимфоцитов в тимусе и приобретению ими соответствующих рецепторов в периферических органах. Местом выработки гормонопо-добных веществ, возможно, служат клетки ретикуло-эпителия и ти-мусные тельца.
При достижении животным половой зрелости тимус подвергается инволютивным изменениям, то есть обратному развитию: специализированная ткань постепенно замещается жировой в большей части долек. В редких случаях при недостатке глюкокортикоидной активности коры надпочечников тимус не претерпевает возрастной инволюции. В этих случаях {statusthymicolymphaticus) у организма отмечают пониженную сопротивляемость инфекциям, и смерть может наступить даже в результате сравнительно легких стрессов. Инволюцию резко ускоряют различные внешние воздействия: стрессы, тяжелые травмы, интоксикации, хронические заболевания, высокая концентрация в крови
 |
глюкокортикоидных гормонов, голодание. При этом наблюдают быст рый выброс из тимуса в общий кровоток Т-лимфоцитов и их активную гибель. Указанные изменения получили название временной, или ащи дентальной, инволюции, которая, в отличие от возрастной, обратима
Врожденная аплазия вилочковой железы у новорожденных живот,, ных сопровождается тяжелыми трофическими и иммунными наруще [ ниями. Этот симптомокомплекс носит название Wasting-синдром и ха! рактеризуется отставанием в росте, истощением, выпадением шерсти дерматитами и диареей. При этом во вторичных органах иммуногенеза отмечают деструкцию лимфоидных узелков и лимфоцитов в них, атрофию лимфоидной ткани и гиперплазию ретикулоэндотелиальных элементов. В периферической крови обнаруживают лимфопению и нейт-рофилез. Иммунные реакции снижены. У молодых животных синдром может развиться и в результате акцидентальной инволюции.
В отношении тимуса на сегодняшний день установлено, что в этом главном органе иммуногенеза активно и непрерывно образуются Т-лимфоциты. Он ответственен за регуляцию иммунного ответа, за пролиферацию и дифференцировку тимусзависимых лимфоцитов в периферических лимфоидных органах путем выделения в кровоток пептидных гормонов (тимозин, тимопоэтин и др.).
Лимфатические узлы. Лимфоузлы (noduslymphaticus) представляют собой овальные или бобовидные структуры, встречающиеся по ходу лимфатических сосудов. У птиц специальные лимфатические органы, подобные лимфоузлам млекопитающих, отсутствуют. У жвачных и некоторых грызунов лимфоузлы располагаются одиночно (моно-нодозный тип), у лошади — часто пакетами или группами (полино-дозный тип). Функция лимфатического аппарата заключается в очищении протекающей лимфы от инородных тел (микроскопических частиц пыли, сажи, микроорганизмов) и в обогащении лимфы эф-фекторными Т- и В-клетками. Таким образом, лимфатические узлы — это важнейшие органы, выполняющие барьерно-фильтрацион-ную функцию.
Лимфатический узел состоит из следующих компонентов: капсулы с трабекулами, разделяющими узел на отсеки; собственно лимфоидной ткани, локализующейся в периферической, промежуточной и центральной частях органа; системы лимфатических синусов и ретикулярных клеток, обеспечивающих нормальное функционирование узла (рис. 8.3).
Капсула (capsula). Она построена из плотной соединительной ткани; в сторону ворот (hilus) от нее отходят трабекулы (trabeculae), фор" мирующие остов органа. Капсула часто связана с жировой клетчаткой, окружающей орган. В трабекулах обнаруживают пучки ретикулярных, коллагеновых, нервных волокон, гладкомышечные клетки, кровеносные сосуды. Сокращение гладких миоцитов способствует прохожие* нию лимфы через узел.
Лимфоидная ткань (textuslymphaticus). Паренхиму органа подразделяют на корковое и мозговое вещество, между которыми выделяют па-Ракортикальную, или тимусзависимую зону. Размеры зон варьируют что обусловлено индивидуальными, видовыми и функциональными особенностями.
Строму узла по морфологии подразделяют на два вида. В лимфоид-ных узелках, входящих в состав коркового вещества и богатых В-лим-фоцитами, преобладают дендритные ретикулярные клетки со светлой цитоплазмой, крупным округлым ядром и множеством выр0С -тов-дендритов. Эти клетки не способны к фагоцитозу, но зато могут адсорбировать на своей поверхности большое количество антигена. Они сохраняют «память» об антигенах и передают ее лимфоцитам.
В паракортикальной зоне, богатой Т-лимфоцитами, часто встречаются клетки с полиморфными ядрами и переплетающимися отростками, а также элементы с типичными признаками макрофагов. Два типа ретикулярных клеток лимфатического узла отличаются не только морфологически, но и цитохимически.
Корковое вещество лимфатического узла (cortex) образовано диффуз-но расположенными лимфоидными клетками, первичными и вторичными лимфоидными узлами {nodulusprimariusetsecundarius). Во вторичных узелках, которые образуются из первичных после встречи с антигеном, по периферии различают корону {corona), состоящую из плотно упакованных многочисленных малых лимфоцитов. В центре узелка расположена более светлая, разреженная область, называемая центром размножения {centrumgerminate), а также светлым, или реактивным, центром. В первичных узелках светлые центры отсутствуют. Центральная зона узелков выглядит светлой вследствие того, что в ней локализуются крупные дендритные ретикулярные клетки, В-лимфо-циты, находящиеся на различных стадиях бласттрансформации, многочисленные макрофаги, а также небольшое количество Т-регулятор-ных лимфоцитов (хелперов и супрессоров). Встречаются здесь и многочисленные фигуры митозов.
В светлых центрах происходит сложная кооперация между иммуно-компетентными клетками. Дендритные клетки с помощью иммуно-глобулиновых рецепторов адсорбируют на своей поверхности антигены, вызвавшие иммунный ответ организма и с помощью комплексов антиген — антитело устанавливают связь с лимфоцитами, вызывая их антигензависимое размножение. Центры размножения формируются только в ответ на попадание антигена в лимфоузел через лимфу. В этом процессе участвуют и типичные макрофаги, которые перерабатывают фагоцитированный антиген до молекулярной формы, способной вызвать пролиферацию и дифференцировку В-лимфоцитов. При первичном ответе на антиген требуется достаточно длительный срок для формирования центров размножения и синтеза антител. При повторном ответе на тот же антиген центры размножения появляются быстрее, а количество В-лимфоцитов возрастает интенсивнее.
Паракортикальная, или тимусзависимая, зона (zonathymodependensseuparacortex) в основном содержит Т-лимфоциты, расположенные диффузно, здесь наблюдают их пролиферацию, бласттрансформацию и превращение в популяции эффекторных и регуляторных клеток (сред и Т-лимфоцитов преобладают Т-киллеры). После удаления или при аплазии тимуса паракортикальная зона лимфатического узла атрофируется, и наоборот, при иммунном ответе по клеточному типу указанная зона значительно увеличивается в размерах (гипертрофируется), для нее характерны сосуды особого типа — посткапиллярные венулы с высоким, кубической формы эндотелием, через которые рециркули-руюшие лимфоциты способны мигрировать из крови в лимфу.
Мозговое вещество (medulla) представлено мякотными тяжами {chordatnedullaris). Это основное место образования плазматических клеток из мигрирующих сюда В-лимфоцитов (после размножения во вторичных узелках коры). Кроме В-лимфоцитов, лимфобластов и зрелых плазматических клеток в мозговом веществе присутствуют также макрофаги. Зрелые плазмоциты располагаются ближе к воротам узла. Их количество при ответе по гуморальному типу резко возрастает.
Система лимфатических синусов. Лимфа циркулирует в узле по системе лимфатических синусов {sinuslimphaticus). Различают субкапсуляр-ный, или краевой, синус {sinussubcapsularis), находящийся между капсулой и узелками коры. В него с выпуклой стороны органа впадает приносящий лимфатический сосуд {vaslymphaticusafferens). От краевого синуса берут начало кортикальные промежуточные синусы (sinuscorticalisperin-odularis), расположенные между узелками и трабекулами и переходящие в промежуточные мозговые синусы (sinusmedullaris), локализующиеся между мякотными тяжами. Указанные синусы соединяются с централь-ным синусом, находящимся возле ворот, а из него лимфа, очищенная от инородных элементов и обогащенная лимфоцитами, впадает в выносящий лимфатический сосуд {vaslymphaticusefferens). Стенки внутренних синусов выстланы прерывистым, лишенным базальной мембраны эндотелием («лимфатический эндотелий»). В синусах встречаются свободные клетки — в основном лимфоциты и макрофаги.
Таким образом, лимфатические узлы помимо фильтрационной и обеззараживающей выполняют еще одну важную функцию: участвуют в иммунологических реакциях, формируя клеточный и гуморальный ответ путем выработки эффекторных клеток и плазмоцитов, синтезирующих иммуноглобулины. Лимфатические узлы весьма чувствительны к воздействию внешних факторов и содержанию кортикостероидов в крови.
Лимфоидные узелки. Эти небольшие неинкапсулированные компактные скопления лимфоцитов часто беспорядочно рассеяны в рыхлой соединительной ткани непосредственно под эпителием. Полагают, что после эпителия они выполняют функцию второго эшело-на барьерной защиты. В указанных структурах лимфоциты, по-видимому, обеспечивают локальную иммунную защиту и, возможно, именно здесь В-лимфоциты приобретают рецепторы, с помощью которых распознают антигены, попадающие из внешней среды. Тесная I связь между эпителием и лимфоидной тканью особенно отчетливо видна в области глотки, носоглотки, миндалин языка и в других местах. Эти образования могут располагаться дискретно — в виде соли-тарных узелков (например, в миндалинах) или аггрегироваться (сливаться) в единую непрерывную массу, образуя глоточное лимфоид-ное кольцо. В дистальном отделе тонкого кишечника (у свиньи и крупного рогатого скота) по всей длине на антимезентериальной стороне располагается структура аггрегированных узелков — полосо-видная пейерова бляшка.
Миндалины развиваются из энтодермы второго глоточного кармана и представляют собой складки слизистой оболочки, нередко связанные с выводными протоками желез. Различают небные миндалины (парные), тубарные и глоточные, которые встречаются у всех домашних животных, язычные — у всех домашних животных за исключением мелких жвачных, и околонадгортанную, которую обнаруживают только у свиньи.
У млекопитающих слизистая оболочка пищеварительного тракта продуцирует треть всей массы лимфоцитов. Выработка последних связана с лимфатическими узелками (nodulilymphatici) слизистой оболочки пищеварительного тракта — солитарными, расположенными в пищеводе, желудке, кишечнике, а также аггрегированными узелками на антимезентериальной стороне тонкой кишки. Периферическая часть лимфатических узелков (в глоточном лимфоидном кольце, узелках кишечника) представлена мелкими лимфоцитами, которые в виде пояска или серпа охватывают центральную светлую зону. В некоторых узелках поясок окружает светлый центр в виде кольца. Светлая зона, или зародышевый центр, представлен главным образом В-лимфоцита-ми различной степени дифференцировки, а также макрофагами и ретикулярными клетками. Часто встречаются фигуры митозов. Лимфоциты здесь размножаются не беспрерывно, а только в ответ на антигенную стимуляцию. Активированные В-лимфоциты после пролиферации в узелке покидают его пределы. Часть из них через стенку посткапиллярных венул мигрирует в кровоток.
Все больше накапливается данных о том, что активированные лимфоциты из лимфоидных узелков могут быть источником плазматических клеток, а некоторые лимфоидные узелки, тесно связанные с эпителием энтодермального происхождения (тонкий кишечник, миндалины, слепая кишка), служат источниками В-лимфоцитов у млекопитающих. Предполагают, что данная система лимфоидной ткани во многом сходна с клоакальной сумкой птиц.
Гемолимфатические узлы. Это особые образования, связанные не с лимфатическими, а с кровеносными сосудами. Расположены преимущественно в грудной полости (у свиньи вдоль грудной аорты), а также в области почечных сосудов. В паренхиме гемолимфатических узлов, которую не подразделяют на кору и мозговое вещество, происходит эритро- и миелопоэз, поэтому узлы красного цвета.
Селезенка. Селезенка (splen) — это непарный, удлиненной формы периферический орган иммуногенеза и кроверазрушения. Селезенка в структурном и функциональном плане более сложно устроена, чем лимфатический узел, хотя в их строении отмечают определенное сходство. Ее объем и масса сильно варьируют в зависимости от кровенаполнения и активности кроветворной функции. В эмбриогенезе селезенка закладывается из мезенхимы и до появления в костном мозге миелоидной ткани участвует в процессах эритро- и гранулоци-топоэза.
Снаружи орган покрыт плотной, гладкой с поверхности капсулой, выстланной мезотелием. Волокнистая соединительная ткань капсулы содержит коллагеновые и эластические волокна, а также многочисленные гладкомышечные клетки. От капсулы внутрь органа отходят многочисленные трабекулы (до 100 на 1 см2 внутренней поверхности), причем с возрастом количество трабекулярной ткани увеличивается. В глубоких слоях селезенки трабекулы анастомозируют между собой и, таким образом, капсула вместе с трабекулами формирует опорно-сократительный аппарат органа.
Межтрабекулярная ткань носит название пульпы (pulpasplenica). В селезенке выделяют красную и белую пульпу (рис. 8.4). Белая пульпа в нативном материале представлена светло-серыми, округлыми, диффузно разбросанными образованиями. Всю остальную часть пространства занимает красная пульпа, где в венозных синусах депонируется кровь, придающая яркокрасный цвет всему органу. Строму селезенки составляет ретикулярная ткань. Соотношение между белой и красной пульпой зависит от кровенаполнения селезенки и функционального состояния ее лимфоидной ткани, а также обусловлено видовыми особенностями. Например, в селезенке лошади преобладает депонирующая функция, поэтому в органе сильно развита трабеку-лярная система, но мало лимфоидной ткани. Масса селезенки лошади составляет 0,375 массы тела, что существенно больше, чем у других животных.
Белая пульпа (pulpaalba) имеет сходное строение с лимфатическими узелками лимфоузлов. Ее лимфоидная ткань формирует шаровидной формы лимфатические узелки, от которых в виде хвостов отходят лимфоидные муфты, или влагалища, окружающие так называемые центральные артерии. Последние проникают в лимфатический узелок, располагаясь в нем чаще эксцентрично, и отдают тонкие коллатерали в его периферический поясок, или краевую зону. По периферии и в центральной части узелков находится сеть грубых ретикулярных волокон (в центре они тоньше и расположены реже, чем на периферии) и сеть эластических волокон, которая тесно связана со стенкой центральной артерии. Лимфатические узелки селезенки, также как и лимфатических узлов, состоят по периферии из мелких Т- и В-лимфоцитов, а более светлая зона — центр размножения — из пролиферирующих

В-лимфоцитов, плазмоцитов, ретикулярных клеток и макрофагов. В периартериальной ткани локализуется, как правило, популяция Т-лимфоцитов. После пролиферации в результате антигенной стимуляции лимфоциты мигрируют в синусы краевой зоны узелка. Через артериальную сеть попадают сюда и В-клетки. Белая пульпа селезенки составляет, %: у лошади 2,5...4, овцы 15...24, крупного рогатого скота 20...21, кролика до 35, у свиньи 11. Однако при достижении половой зрелости объем белой пульпы у всех животных снижается.
Красная пульпа (pulparubra) занимает пространство между белой пульпой и трабекулами. Она состоит из трехмерной сети ретикулярных волокон и многочисленных венозных синусоидов с диаметром просвета от 12 до 40 мкм, пронизывающих сеть. Эндотелиальные клетки сосудов палочковидной формы и между ними обнаруживают множество щелевидных пространств, хорошо контурирующих на поперечных срезах. Вокруг синусоидов проходят толстые кольцевые ре-
248 тикулярные волокна, охватывающие их стенки наподобие «обручей». Характерная особенность красной пульпы — наличие между синусоидами большого количества форменных элементов крови, особенно эритроцитов, придающих ей красноватый цвет, а также гранулоцитов и макрофагов. Синусы селезенки и межсосудистая ретикулярная ткань, составляющие красную пульпу, могут депонировать в зависимости от вида животных от 10...15 % крови (у кошки, собаки; у лошади существенно больше).
Васкуляризация и иннервация селезенки. Место вхождения кровеносных сосудов и нервов на внутренней поверхности селезенки называется воротами. Трабекулярные артерии (arteriatrabeculars) после выхода из трабекул переходят в пульпарные (arteriapulpaerubra), которые следуют в лимфоидную ткань. На некотором расстоянии в их адвентиции появляются периартериальные лимфатические влагалища (vaginaperiarterialislymphatica) и лимфатические узелки (lymphano-dulussplenicus). Артерия, проходящая через узелок, называется центральной (arterialymphonoduli). Сразу же после выхода из узелка она разделяется на несколько мелких артериол, приобретая вид «кисточки» (arteriolapenicillaris). Стенки кисточковых артериол окружены «муфтами», или «гильзами», из скоплений ретикулярных клеток, образующих своеобразные сфинктеры, сужающие или расширяющие эти сосуды. Далее артериолы переходят в капилляры, которые впадают в систему расширенных синусоидов. Затем кровь из красной пульпы направляется в собирательные венулы, из которых попадает в селезеночную вену, выходящую в области ворот. Установлено, что на конце не только артериол, но и на конце синусоида есть сфинктер. Циркуляция крови в органе регулируется сокращением или расслаблением сфинктеров.
Таким образом, синусоиды представляют собой начальный отдел венозной системы селезенки. Форменные элементы крови могут активно проходить через многочисленные щели между ретикулоэндоте-лиальными клетками стенки синусоидов. Предполагается, что состарившиеся или дефектные эритроциты, утратившие эластичность, при этом повреждаются и тотчас фагоцитируются присутствующими в петлях ретикулярной сети макрофагами. Другие же активные и жизнеспособные клетки легко проходят через эти щели. Циклические изменения работы синусоидов определяются артериальными и венозными сфинктерами. При открытии синусов кровь свободно поступает через синусы в вены. Сокращение только венозных сфинктеров приводит к отфильтровыванию плазмы от эритроцитов и кровенаполнению ' красной пульпы. При закрытии тех и других сфинктеров кровь депонируется на определенное время в синусах. В это время состарившиеся эритроциты разрушаются и фагоцитируются, а плазма крови пополняется минеральными и органическими веществами. Стенки синусов и вен состоят из видоизмененного эндотелия.
Вместе с селезеночной артерией через ворота в паренхиму селезенки вступают нервные стволики. В их составе различают миелиновые и безмиелиновые нервные волокна. Первые принадлежат нейронам чувствительных ганглиев, терминальные веточки которых образуют свободные нервные окончания в капсуле, трабекулах, на стенках тра-бекулярных артерий и вен, вблизи синусоидов. Безмиелиновые волокна представлены постганглионарными симпатическими и парасимпатическими эфферентными аксонами, обеспечивающими нейротрофи-ческий и регуляторный контроль микроциркуляторного русла селезенки.
Функциональное значение селезенки. В эмбриогенезе селезенка выполняет функцию кроветворного органа, в постнатальном периоде — органа иммуногенеза: в ней протекает лимфоцитопоэз. В отличие от лимфатических узлов, антитела, образующиеся в селезенке, обеззараживают кровь. В этой связи орган служит своеобразным биологическим фильтром для артериальной крови. Благодаря сложной системе кровоснабжения — наличию многочисленных синусоидов, артериальных и венозных сфинктеров, анастомозов — в селезенке может депонироваться значительное количество крови. В результате периодических сокращений она обеспечивает подачу крови в общее кровяное русло, несколько повышая кровяное давление. В результате гемолиза в красной пульпе происходит постоянная гибель отслуживших свой срок эритроцитов и тромбоцитов, поэтому ее нередко называют «кладбищем» эритроцитов. При этом большая часть высвободившегося из распавшегося гемоглобина железа реутилизируется и используется для формирования новых эритроцитов в красном костном мозге, а билли-рубин — для построения желчи.