| Скачать .docx | Скачать .pdf |
Реферат: Электрофорез 2
Содержание
Принцип метода электрофореза . 3
Физико-химические свойства белков и методы их выделения . 5
Методы выделения и очистки белков. 6
Методы разрушения тканей и экстракции белков. 6
Гомогенизация биологического материала . 6
Метод замораживания и оттаивания ткани . 6
Экстракция белков, связанных с мембранами, и разрушение олигомерных белков на протомеры .. 7
Удаление из раствора небелковых веществ . 7
Очистка белков избирательной денатурацией . 8
Гель-фильтрация, или метод молекулярных сит .. 9
Ионообменная хроматография . 12
Аффинная хроматография, или хроматография по сродству . 14
Очистка белков от низкомолекулярных примесей . 15
MultiNA - Система электрофореза на микрочипе для исследования нуклеиновых кислот. 16
Основные преимущества системы электрофореза на микрочипе: . 17
Введение
Электрофорез занимает сейчас центральное место среди методов исследования белков и нуклеиновых кислот. В современной научной литературе редко можно встретить статью, в которой бы на той или иной стадии фракционирования или характеристики этих биополимеров не был использован электрофорез. Метод позволяет разделять макромолекулы, различающиеся по таким важнейшим параметрам, как размеры (или молекулярная масса), пространственная конфигурация, вторичная структура и электрический заряд, причем эти параметры могут выступать как порознь, так и в совокупности.
Физический принцип метода заключается в следующем. Находящиеся в буферном растворе макромолекулы обладают некоторым суммарным электрическим зарядом, величина и знак которого зависят от рН среды. Если через этот раствор, заключенный в канал из изолирующего материала, например, стеклянную трубку, начать пропускать электрический ток, то вдоль канала установится определенный градиент напряжения, т. е. сформируется электрическое поле. Его напряженность измеряется разностью потенциалов по концам рабочего канала (или его участка), отнесенной к его длине (В/см). Под действием поля макромолекулы в соответствии со своим суммарным зарядом мигрируют в направлении катода или анода, причем их трение об окружающую среду ограничивает скорость миграции. В зависимости от величины заряда и размеров молекулы приобретают разные скорости, и в этом — сущность процесса электрофореза. Постепенно исходный препарат, состоявший из различных молекул, разделяется на зоны одинаковых молекул, мигрирующих с одной и той же скоростью. Со временем эти зоны распределяются по длине канала. В настоящее время почти исключительно используются полиакриламидные гели (ПААГ) и гели агарозы. Варьируя концентрацию полимера, можно получать гели с очень широким диапазоном размеров пор. Кроме того, можно изменять электрические заряды макромолекул путем вариации рН буфера, а их конфигурацию путем введения в буфер денатурирующих агентов или детергентов. Все это придает методу электрофореза исключительную гибкость.
В ходе электрофореза зоны растворенных макромолекул остаются невидимыми. Для наблюдения за процессом в исходный препарат добавляют краситель, молекулы которого несут электрический заряд того же знака, что и фракционируемые макромолекулы, но не взаимодействуют с ними. Краситель тоже передвигается в электрическом поле, но уже в виде окрашенной зоны. Его подбирают таким образом, чтобы скорость миграции наиболее подвижных макромолекул была несколько ниже, чем у молекул красителя. Когда окрашенная зона доходит до конца трубки, электрофорез прекращают.
Разделившиеся зоны биополимеров во избежание их диффузии немедленно фиксируют. Для этого гель извлекают из стеклянной формы и вымачивают в смеси кислоты со спиртом так, что белки или нуклеиновые кислоты выпадают в осадок в том самом месте, где закончилась их миграция в ходе электрофореза. После фиксации (или одновременно с ней) проводят окрашивание зон путем вымачивания геля в растворе красителя, прочно связывающегося с белком или нуклеиновой кислотой. Излишек красителя удаляют.
Вместо цилиндрических часто используют гели в виде тонких пластин, заполимеризованные между двумя плоскими стеклами. Такие пластины имеют важное преимущество: на них можно одновременно фракционировать несколько препаратов. Обычно их вносят с одного края геля на равных расстояниях друг от друга. Каждый препарат разделяется в электрическом поле независимо от своих соседей, образуя свой набор зон. Вместо окрашивания или наряду с ним часто используют методы обнаружения разделенных зон по их радиоактивности. К ним относятся приемы регистрации полос на фотопленке посредством авторадиографии или флюорографии и различные способы счета радиоактивности в геле с помощью жидкостных сцинтилляционных счетчиков.
Фракционированием в ПААГ и агарозе не исчерпываются современные методы электрофореза. В качестве «носителей» жидкой фазы широко используют также пленки из ацетата целлюлозы, фильтровальную бумагу, тонкие слои силикагеля, целлюлозы, сефадекса и др. В некоторых случаях, например для разделения низкомолекулярных веществ, эти системы имеют свои преимущества, однако для фракционирования белков, нуклеиновых кислот и их фрагментов в настоящее время используют почти исключительно гель-электрофорез
Физико-химические свойства белков и методы их выделения
Важное место в биохимических исследованиях занимает выделение индивидуальных белков из органов и тканей. Очищенные индивидуальные белки нужны для изучения их первичной структуры, получения кристаллов белков с целью исследования их пространственной структуры методом рентгеноструктурного анализа, установления взаимосвязи между первичной, пространственной структурой белка и его функцией.
Некоторые очищенные индивидуальные белки используют в медицине как лекарственные препараты, например гормон инсулин применяют для лечения сахарного диабета, а пищеварительные ферменты поджелудочной железы назначают при нарушении её функций в качестве заместительной терапии. Кроме того, очищенные ферменты часто используют в биохимических исследованиях в качестве химических реактивов для определения веществ в биологических жидкостях.
Большинство методов, используемых для очистки индивидуальных белков, основано на различиях их физико-химических свойств, а также возможности специфично связываться с лигандом.
Методы выделения и очистки белков
Получение индивидуальных белков из биологического материала (тканей, органов, клеточных культур) требует проведения последовательных операций, включающих:
· дробление биологического материала и разрушение клеточных мембран;
· фракционирование органелл, содержащих те или иные белки;
· экстракцию белков (перевод их в растворённое состояние);
· разделение смеси белков на индивидуальные белки.
Методы разрушения тканей и экстракции белков
Для разрушения биологического материала используют методы: гомогенизации ткани, метод попеременного замораживания и оттаивания, а также обработку клеток ультразвуком.
Гомогенизация биологического материала
Ткань, находящуюся в буферном растворе с определённым значением рН и концентрацией солей, помещают в стеклянный сосуд (гомогенизатор) с пестиком. Вращающийся пестик измельчает и растирает ткань о притёртые стенки сосуда.
Метод замораживания и оттаивания ткани
В результате попеременного замораживания и оттаивания образующиеся кристаллы льда разрушают оболочки клеток.
После разрушения ткани нерастворимые части осаждают центрифугированием. Последующее центрифугирование гомогената с разной скоростью позволяет получить отдельные фракции, содержащие клеточные ядра, митохондрии и другие органеллы, а также надосадочную жидкость, в которой находятся растворимые белки цитозоля клетки. Искомый белок будет содержаться в одной из этих фракций.
Экстракция белков, связанных с мембранами, и разрушение олигомерных белков на протомеры
Если искомый белок прочно связан с какими-либо структурами клетки, его необходимо перевести в раствор. Так, для разрушения гидрофобных взаимодействий между белками и липидами мембран в раствор добавляют детергенты; чаще всего используют тритон Х-100 или додецилсульфат натрия.
При действии детергентов обычно разрушаются и гидрофобные взаимодействия между протомерами в олигомерных белках.
Удаление из раствора небелковых веществ
Нуклеиновые кислоты, липиды и другие небелковые вещества можно удалить из раствора, используя их Особенные физико-химические свойства. Так, липиды легко удаляются из раствора добавлением органических растворителей, например ацетона. Однако воздействие должно быть кратковременным, так как ацетон вызывает денатурацию некоторых белков. Нуклеиновые кислоты осаждают добавлением в раствор стрептомицина.
Методы очистки белков
Наиболее трудоёмкий этап получения индивидуальных белков - их очистка от других белков, находящихся в растворе, полученном из данной ткани. Часто изучаемый белок присутствует в небольших количествах, составляющих доли процента от всех белков раствора.
Так как белки обладают конформационной лабильностью, при работе с белками следует избегать денатурирующих воздействий, поэтому выделение и очистка белков происходят при низких температурах.
На первых стадиях очистки белков целесообразно использовать методы, учитывающие какую-либо характерную особенность данного белка, например термостабильность или устойчивость в кислых растворах. Первыми методами очистки необходимо удалить из раствора основную массу балластных белков, которые значительно отличаются от выделяемого белка физико-химическими свойствами. Впоследствии применяют всё более тонкие методы очистки белка.
Очистка белков избирательной денатурацией
Большинство белков денатурирует и выпадает в осадок уже при кратковременном нагревании раствора до 50-70°С или подкислении раствора до рН 5. Если выделяемый белок выдерживает эти условия, то с помощью избирательной денатурации можно удалить большую часть посторонних белков, отфильтровав выпавшие в осадок-белки, или осадить их центрифугированием.
Высаливание
Метод очистки белков, основанный на различиях в их растворимости при разной концентрации соли в растворе. Соли щелочных и щёлочно-земельных металлов вызывают обратимое осаждение белков, т.е. после их удаления белки вновь приобретают способность растворяться, сохраняя при этом свои нативные свойства.
Чаще всего для разделения белков методом высаливания используют разные концентрации солей сульфата аммония - (NH4 )2 SO4 . Чем выше растворимость белка, тем большая концентрация соли необходима для его высаливания.
Гель-фильтрация, или метод молекулярных сит
Для разделения белков часто используют хроматографические методы, основанные на распределении веществ между двумя фазами, одна из которых подвижная, а другая неподвижная. В основу хроматографических методов положены разные принципы: гель-фильтрации, ионного обмена, адсорбции, биологического сродства.
Метод разделения белков с помощью гель-фильтрационной хроматографии основан на том, что вещества, отличающиеся молекулярной массой, по-разному распределяются между неподвижной и подвижной фазами. Хроматографическая колонка заполняется гранулами пористого вещества (сефадекс, агароза и др.). В структуре полисахарида образуются поперечные связи и формируются гранулы с "порами", через которые легко проходят вода и низкомолекулярные вещества. В зависимости от условий можно формировать гранулы с разной величиной "пор".
Неподвижная фаза - жидкость внутри гранул, в которую способны проникать низкомолекулярные вещества и белки с небольшой молекулярной массой. Смесь белков, нанесённую на хроматографическую колонку, вымывают (элюируют), пропуская через колонку растворитель. Вместе с фронтом растворителя движутся и самые крупные молекулы.
Более мелкие молекулы диффундируют внутрь гранул сефадекса и на некоторое время попадают в неподвижную фазу, в результате чего их движение задерживается. Величина пор определяет размер молекул, способных проникать внутрь гранул .
Так как гелевая структура сефадекса легко деформируется под давлением, гели стали заменять более жёсткими матрицами (сефактил, той-оперл), представляющими сферические гранулы с разными размерами пор. Выбор размеров пор в гранулах зависит от целей хроматографии (о других хроматографических методах будет сказано ниже).
Ультрацентрифугирование
Метод разделения также основан на различии в молекулярных массах белков. Скорость седиментации веществ в процессе вращения в ультрацентрифуге, где центробежное ускорение достигает 100 000-500 000 g, пропорционально их молекулярной массе. На поверхность буферного раствора, помещённого в кювету, наносят тонкий слой смеси белков. Кювету помещают в ротор ультрацентрифуги. При вращении ротора в течение 10-12 ч более крупные молекулы (с большей молекулярной массой) оседают в буферном растворе с большей скоростью. В результате в кювете происходит расслоение смеси белков на отдельные фракции с разной молекулярной массой .После расслоения белковых фракций дно кюветы прокаливают иглой и по каплям собирают содержимое небольшими порциями в пробирки.
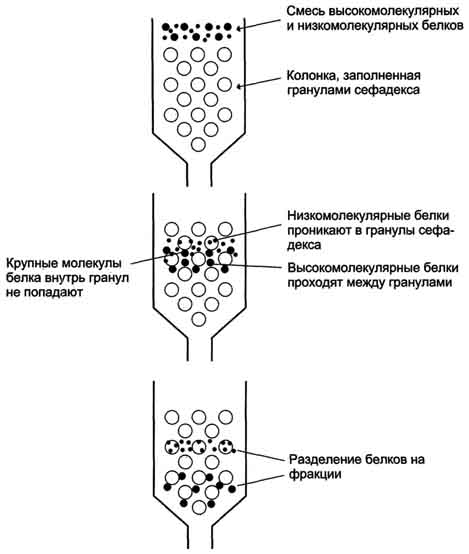
Разделение смеси белков методом гель-фильтрации.
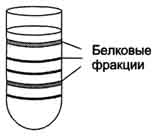
Кювета, заполненная буферным раствором с разделёнными белковыми фракциями.
Электрофорез белков
Метод основан на том, что при определённом значении рН и ионной силы раствора белки двигаются в электрическом поле со скоростью, пропорциональной их суммарному заряду. Белки, имеющие суммарный отрицательный заряд, двигаются к аноду (+), а положительно заряженные белки - к катоду (-).
Электрофорез проводят на различных носителях: бумаге, крахмальном геле, полиакриламидном геле и др. В отличие от электрофореза на бумаге, где скорость движения белков пропорциональна только их суммарному заряду, в полиакриламидном геле скорость движения белков пропорциональна их молекулярным массам.
Разрешающая способность электрофореза в полиакриламидном геле выше, чем на бумаге. Так, при электрофорезе белков сыворотки крови человека на бумаге обнаруживают только 5 главных фракций: альбумины, α1 глобулины, α2 -глобулины, β-глобулины и γ-глобулины (рис. 1-57). Электрофорез тех же белков в полиакриламидном геле позволяет получить до 18 различных фракций. Для обнаружения белковых фракций полоски бумаги или столбики геля обрабатывают красителем (чаще всего бромфеноловым синим или амидовым чёрным). Окрашенный комплекс белков с красителем выявляет расположение различных фракций на носителе.
Ионообменная хроматография
Так же как и электрофорез, метод основан на разделении белков, различающихся суммарным зарядом при определённых значениях рН и ионной силы раствора. При пропускании раствора белков через хроматографическую колонку, заполненную твёрдым пористым заряженным материалом, часть белков задерживается на нём в результате электростатических взаимодействий.
В качестве неподвижной фазы используют ионообменники - полимерные органические вещества, содержащие заряженные функциональные группы.
Различают положительно заряженные анионообменники, среди которых наиболее часто используют диэтиламиноэтилцеллюлозу (ДЭАЭ-целлюлозу), содержащую катионные группы, и отрицательно заряженные катионообменники, например карбоксиметилцеллюлозу (КМ-цел-люлозу), содержащую анионные группы.
![]()
Выбор ионообменника определяется зарядом выделяемого белка. Так, для выделения отрицательно
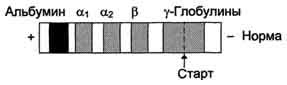
Электрофорез белков сыворотки крови здорового человека на бумаге.
заряженного белка используют анионообменник. При пропускании раствора белка через колонку прочность связывания белка с анионообменником зависит от количества отрицательно заряженных карбоксильных групп в молекуле. Белки, адсорбированные на анионообменнике, можно смыть (элюировать) буферными растворами с различной концентрацией соли, чаще всего NaCI, и разными значениями рН. Ионы хлора связываются с положительно заряженными функциональными группами анионообменника и вытесняют карбоксильные группы белков. При низких концентрациях соли элюируются белки, слабо связанные с анионообменником. Постепенное увеличение концентрации соли или изменение рН, что меняет заряд белковой молекулы, приводит к выделению белковых фракций, в одной из которых находится искомый белок.
Аффинная хроматография, или хроматография по сродству
Это наиболее специфичный метод выделения индивидуальных белков, основанный на избирательном взаимодействии белков с лигандами, прикреплёнными (иммобилизированными) к твёрдому носителю. В качестве лиганда может быть использован субстрат или кофермент, если выделяют какой-либо фермент, антигены для выделения антител и т.д. Через колонку, заполненную иммобилизованным лигандом, пропускают раствор, содержащий смесь белков. К ли-ганду присоединяется только белок, специфично взаимодействующий с ним; все остальные белки выходят с элюатом. Белок, адсорбированный на колонке, можно снять, промыв её раствором с изменённым значением рН или изменённой ионной силой. В некоторых случаях используют раствор детергента, разрывающий гидрофобные связи между белком и лигандом.
Аффинная хроматография отличается высокой избирательностью и помогает очистить выделяемый белок в тысячи раз. 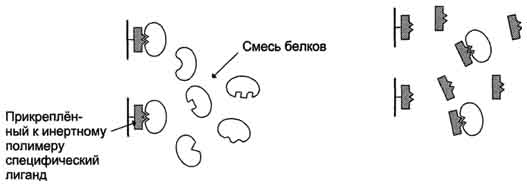
Аффинная хроматография.
Очистка белков от низкомолекулярных примесей
Для удаления низкомолекулярных соединений, в частности сульфата аммония после высаливания, применяют диализ. Метод основан на том, что через полупроницаемую мембрану, пропускающую низкомолекулярные вещества, не проходят белки, имеющие более высокую молекулярную массу. В стакан большой ёмкости (около 1 л) с буферным раствором помещают полупроницаемый мешочек, заполненный раствором белка с солью.
Скорость выхода соли из мешочка в буферный раствор пропорциональна градиенту его концентраций по обе стороны от мембраны. По мере выхода соли из мешочка буферный раствор в стакане меняют.
Для очистки белков от низкомолекулярных примесей используют также метод гель-фильтрации.
Для определения частоты (гомогенности) выделенного белка применяют методы с высокой разрешающей способностью, например электрофорез в полиакриламидном геле, высокоэффективная хроматография высокого давления. От чистоты лекарственного белкового препарата зависят его биологическая эффективность и аллергенность (т.е. способность вызывать аллергические реакции). Чем качественнее очищен препарат, тем меньше вероятность осложнений при его применении.
Нуклеиновые кислоты - это биополимеры, макромолекулы которых состоят из многократно повторяющихся звеньев - нуклеотидов. Поэтому их называют также полинуклеотидами. Важнейшей характеристикой нуклеиновых кислот является их нуклеотидный состав. В состав нуклеотида - структурного звена нуклеиновых кислот - входят три составные части:
· азотистое основание - пиримидиновое или пуриновое. В нуклеиновых кислотах содержатся основания 4-х разных видов: два из них относятся к классу пуринов и два - к классу пиримидинов. Азот, содержащийся в кольцах, придает молекулам основные свойства.
· моносахарид - рибоза или 2-дезоксирибоза. Сахар, входящий в состав нуклеотида, содержит пять углеродных атомов, т.е. представляет собой пентозу. В зависимости от вида пентозы, присутствующей в нуклеотиде, различают два вида нуклеиновых кислот - рибонуклеиновые кислоты (РНК), которые содержат рибозу, и дезоксирибонуклеиновые кислоты (ДНК), содержащие дизоксирибозу.
· остаток фосфорной кислоты. Нуклеиновые кислоты являются кислотами потому, что в их молекулах содержится фосфорная кислота. Нуклеотид - фосфорный эфир нуклеозида. В состав нуклеозида входят два компонента: моносахарид (рибоза или дезоксирибоза) и азотистое основание.
MultiNA - Система электрофореза на микрочипе для исследования нуклеиновых кислот
Сочетание технологии электрофореза на микрочипе, автоматизированной пробоподготовки и чувствительного флуориметрического детектирования предлагает высокоэффективную замену традиционному агарозному гель-электрофорезу. Качественный и количественный анализ нуклеиновых кислот становится теперь как никогда быстрым, недорогим и высокоточным. В настоящее время при проведении исследований нуклеиновых кислот основным методом изучения распределения фрагментов ДНК и РНК по молекулярной массе (размеру) остается агарозный гель-электрофорез. К основным недостаткам этого метода следует отнести достаточно высокую стоимость анализа при использовании готовых пластин геля, высокую трудоемкость, связанную с большим количеством ручных операций, длительность анализа, необходимость использования дополнительного весьма дорогостоящего оборудования для визуализации и последующей цифровой обработки электрофореграмм, использование небезопасных для здоровья реагентов (бромистого этидия). Всех этих недостатков лишена последняя разработка компании Shimadzu – прибор для электрофоретического разделения нуклеиновых кислот с использованием микрочипа MCE®-202 MultiNA.
Основные преимущества системы электрофореза на микрочипе:
- Низкая стоимость анализа. Конструкция и материал микрочипа позволяют использовать его для нескольких тысяч анализов, при этом используется крайне незначительное количество расходных материалов. Тем самым достигается существенное снижение стоимости (в 1,5 – 3 раза) одного анализа ДНК по сравнению с агарозным гель-электрофорезом. Если сравнивать систему электрофореза на микрочипе с существующими сейчас на рынке системами капиллярного электрофореза, то снижение стоимости анализа может достигать 6 и более раз. В случае анализа РНК экономия может быть еще более существенной (рассчитано, исходя из стоимости оборудования и реагентов на японском рынке).
- Высокая скорость анализа. Высокоскоростной автоматизированный анализ до 108 образцов (96 + 12 дополнительно). С использованием одного микрочипа полный цикл анализа ДНК составляет 255 секунд. Для увеличения производительности в прибор может быть установлено до четырех микрочипов для параллельной работы. В этом случае время одного анализа сокращается до 75 секунд.
- Высокая чувствительность. Флуориметрический детектор с фотоумножителем и источником возбуждения флуоресценции на основе светодиода обеспечивает примерно 10-кратное увеличение чувствительности по сравнению с традиционным окрашиванием бромистым этидием (данные получены в ходе внутренних испытаний в лаборатории Shimadzu). К тому же используемые флуоресцентные красители, такие как SYBR green II и SYBR gold, абсолютно безвредны для здоровья в отличие от бромистого этидия.
- Высокое разрешение и прекрасная воспроизводимость анализа. Оптимальная конфигурация капилляров микрочипа и специально подобранный состав буферного раствора обеспечивают превосходные характеристики электрофоретического разделения нуклеиновых кислот. Благодаря автоматизированной системе пробоподготовки количество ручных операций сведено к минимуму. Внутренние маркеры молекулярного веса уже включены в наборы реагентов и используются при каждом анализе. Все это в комплексе существенно увеличивает надежность и воспроизводимость получаемых результатов. Прибор может комплектоваться четырьмя различными наборами реагентов для анализа ДНК разного размера и РНК
- Простота использования. Программное обеспечение с дружественным пользовательским интерфейсом и большим количеством функций максимально упрощает проведение исследования. Оператору достаточно загрузить образцы и реагенты в прибор, задать в программе желаемую последовательность анализа образцов и кликнуть по иконке «Старт» на экране компьютера. Все остальное, включая обработку электрофореграмм, прибор выполнит в автоматическом режиме.
Микрочип MultiNA
Микрочип, изготовленный из кварца высокой чистоты, включает микроемкости для загрузки образца и реагентов и электрофоретический канал 23 × 0,09 × 0,05 мм (д ×ш × г). Напряжение подается при помощи напыленных платиновых электродов. Канал и микроемкости выполнены с высочайшей точностью при помощи уникальной фотолитографической технологии Shimadzu. Специальное покрытие обеспечивает возможность многократного использования одного и того же микрочипа (~ 3600 анализов). Следствием малой длины электрофоретического канала и его оптимальной формы является непревзойденная на сегодняшний день скорость электрофоретического разделения нуклеиновых кислот. Наборы реагентов для анализа ДНК и РНК. Прибор может комплектоваться тремя наборами реагентов для анализа ДНК различного размера и набором реагентов для анализа РНК.