| Скачать .docx |
Реферат: В. Д. Кабанов теория высокой скорости роста свиней и использование ее в селекции
В.Д. Кабанов
ТЕОРИЯ ВЫСОКОЙ СКОРОСТИ РОСТА СВИНЕЙ И ИСПОЛЬЗОВАНИЕ ЕЕ В СЕЛЕКЦИИ
Проблемная лекция
В лекции изложены пути и методы повышения скорости роста свиней на интенсивном откорме до 900-1000 г в сутки при условии интенсивного кормления животных по современным нормам (2003).
Рецензенты: В.В. Горай, доктор сельскохозяйственных наук, Н.М. Костомахин, доктор сельскохозяйственных наук, профессор.
СОДЕРЖАНИЕ
Введение …...…………………………………………………………………………………… 3
1. Критерии и методы учета роста …………………………………………………….……. 4
2. Механизмы и ресурсы повышения скорости роста ………………………..………….. 7
3. Наследственность и изменчивость признаков роста ………………………………. 20
4. Использование признаков роста в селекции
в целях повышения скорости роста свиней ………………………………………… 21
5. Разработка методов управления процессом роста и развития в онтогенезе ……... 23
5.1. Моделирование процесса роста ……………………………………………… 23
5.2. Создание оптимальной модели роста
для скороспелой мясной породы СМ-1 ……………………………………… 24
5.3. Практическая реализация теории ……………………………………………. 26
Заключение …………………………………………………………………………………... 28
Библиографический список ………………………………………………………………... 29
Введение
Проблема роста принадлежит к числу наиболее важных проблем в биологии. Очевидно не будет ошибкой сказать, что значение роста в широком смысле слова состоит в том, что он является единственным условием реализации наследственности в процессе роста и дифференциации клеток, в результате которого каждая дочерняя клетка получает такой же, как у материнской клетки, набор хромосом, что происходит при митозе. Механизм реализации наследственности хорошо прослеживается на примере синтеза ДНК, свойственного, по выражению К.Г. Ларка (16), процессу размножения, а не всему широкому кругу явлений, обычно связанных с активно функционирующими клетками. Он говорит, что нередко в клетках, где процессы клеточного метаболизма протекают весьма активно, синтез ДНК оказывается незначительным или не идет вовсе. Клеточному делению, напротив, обязательно предшествует репликация генетического материала, без которого невозможно образование полноценных дочерних клеток… В нормальном цикле клеточного деления содержание ДНК в клетке, по данным как химических, так и генетических измерений, точно удваивается. Клетки, в которых синтез ДНК и деление были временно приостановлены, обычно сначала возобновляют синтез ДНК, а затем уже начинают делиться.
Значение роста как непрерывного саморегулируемого процесса в узком смысле определяется единственной в своем роде способностью растений и животных наращивать биомассу. В оценке его роли применительно к животноводству мы не можем не согласиться с мнением американского профессора У. Дэвинпорта, который еще в начале прошлого столетия говорили, что важность изучения процесса роста и влияния на него различных факторов никоим образом не может быть переоценена и тот факт, что почти не поддающиеся исчислению громадные количества мяса…. ежегодно вновь образуются, благодаря процессу роста, достаточен для того, чтобы проблема роста во всей ее широте заняла в биологии главенствующее положение.
Однако с развитием генетики проблема роста была незаслуженно отодвинута на задний план, где из поля зрения большинства исследователей выпало значение роста как селекционного признака, а основное внимание переключалось на изучение его механизма, хотя и встречаются работы, в которых вопросы морфогенеза рассматриваются в связи с генетикой (28,24). Но и в них еще не получены ответы на многие вопросы роста. По признанию К. Уоддингтона, наука не может претендовать на всеобъемлющее понимание генетической детерминации развития до тех пор, пока не будет установлено, каким образом синтезируемые в клетках высших организмов белки координируются количественно и качественно так, что клетка приобретает определенные гистолические свойства.
Такая оценка состояния изученности роста в равной, а, может быть, и значительно большей степени, относится к прикладным исследованиям зоотехнической науки, призванным служить решению больших практических задач в животноводстве. В связи с этим изложенные в данной публикации основные положения теории высокой скорости роста свидетельствуют об эффективности использования ее в селекции и открывают большие возможности по регулированию и управлению процессом роста в онтогенезе и филогенезе, дальнейшему повышению откормочной и мясной продуктивности свиней.
Основные положения теории высокой скорости роста были разработаны в 70-х годах прошлого столетия (6,7), реализованы в процессе создания скороспелой мясной породы свиней (8,9). Она предусматривает повышение скорости роста свиней до 900-1000 г в сутки при условии полноценного кормления животных по современным нормам (2003 г.).
В разработку теории входит:
- определение критериев и методов учета роста;
- изучение механизма и ресурсов повышения скорости роста;
- изучение наследственности и изменчивости отдельных элементов и закономерностей роста для использования их в селекции;
- разработка методов управления процессом роста и развития в онтогенезе и филогенезе.
1. Критерии и методы учета роста
Процесс роста мы рассматриваем как приращение массы тела во времени. Скорость роста – величина приращения массы тела в единицу времени.
Наиболее простым и доступным выражением роста можно считать массу тела животного в определенном возрасте, а прироста – единицу массы прироста в единицу времени.
![]() где,
где,
V1 и V2 – масса тела в начале и конце учитываемого периода, а t1 и t2 – время в начале и конце учитываемого периода.
В поисках надежной характеристики процесса постэмбрионального роста мы исходим из модели, принятой Шмальгаузеном (28) в виде формулы, характеризующей пораболу:
![]()
где m – постоянная величина, характеризующая начальную массу; k – константа роста, выражающая интенсивность этого процесса.
Если принять скорость роста как величину бесконечно малых приращений величин массы в бесконечно малое время, то константу роста можно найти путем последовательного вычисления уравнений:
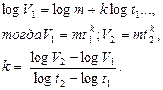
Учитывая, что скорость роста и дифференциация находятся в обратном соотношении друг к другу, дается вторая формула для вычисления константы (28):
![]()
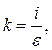
где i
– скорость роста индифферентных тканей; ![]() - скорость дифференцировки. Делением константы на возраст получаем выражение скорости роста в любом возрасте.
- скорость дифференцировки. Делением константы на возраст получаем выражение скорости роста в любом возрасте.
Однако рассматриваемый способ выражения скорости роста основан на допущении равномерности прироста во времени. Из-за неравномерности постэмбрионального роста высших животных использование констант можно считать недостаточным. Это в равной степени относится к методу составления теоретических уравнений асимптотической кривой, использованному (21) для описания нисходящей ветви S-образной кривой, отражающей рост крупного рогатого скота, выраженный формулой:
![]()
![]() где
где
А
– максимальное значение признака, к которому стремится весь ряд, т.е. средняя живая масса одного пола данной породы; ![]() - сумма прироста в постэмбриональный период;
- сумма прироста в постэмбриональный период; ![]() - приращение логарифма разности между максимальным и текущим значениями прироста в единицу времени,
- приращение логарифма разности между максимальным и текущим значениями прироста в единицу времени, ![]() - возраст, выраженный в любых интервалах.
- возраст, выраженный в любых интервалах.
Использование рассмотренных критериев (начальная масса, константа и длительность течения роста) больше подходят для выражения лишь общей модели роста и недостаточно для характеристики, а тем более познания механизма и прогнозирования процесса роста в онтогенезе, столь необходимых для использования признаков роста в селекции. Уместно будет признать, что вряд ли такой сложный процесс, каким представляется рост в его многообразной зависимости от внешних и внутренних факторов, можно выразить формулой. Важнее, по-видимому, полнее изучить отдельные составные части этого взаимосвязанного и взаимообусловленного процесса.
В целях более полной характеристики внутривидовых (породных, половых, индивидуальных) особенностей роста свиней мы предлагаем рассматривать следующие его элементы: 1) скорость, 2) интенсивность, 3) продолжительность, 4) великорослость и 5) скороспелость великорослости.
Скорость роста – это прирост тела в единицу времени. Она является абсолютной мерой роста за период, в который измеряется, и выражается в единицах прироста за единицу времени, например, граммах.
Скорость постэмбрионального роста, выраженная, например в граммах, представляет собой отношение разности живой массы животного в момент измерения (
V)
и массой тела при рождении![]() к его возрасту (t), выраженному в сутках.
к его возрасту (t), выраженному в сутках.
![]()
На отдельных этапах онтогенеза ![]() измеряется величиной прироста в единицу времени (темп роста):
измеряется величиной прироста в единицу времени (темп роста):
![]()
Хозяйственными показателями скорости роста могут служить живая масса животных в определенном возрасте и возраст, в котором они достигают определенной живой массы.
Интенсивность роста – показатель степени напряженности этого процесса, представляет собой относительную скорость роста и выражается в процентах к показателю, с которым соотносится.
В биологии и зоотехнии широкое распространение получило определение коэффициента роста (27)
![]()
показывающего отношение взрослого животного к его живой массе при рождении.
Для сравнительной оценки относительной скорости роста животных разной живой массы, не потеряла своего значения формула (17), показывающая отношение прироста за определенный период к начальной массе (величине) организма, выраженное в процентах:
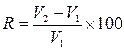
Эта формула в модификации С. Боди (1943) имеет следующий вид:

где R
– относительный прирост (коэффициент относительной скорости роста); ![]() - начальная живая масса;
- начальная живая масса; ![]() - живая масса в конце периода. В данном случае отношение прироста определяется не к начальной, а к средней величине животного.
- живая масса в конце периода. В данном случае отношение прироста определяется не к начальной, а к средней величине животного.
Интенсивность роста более наглядно обозначать символом ![]() и выражать отношение живой массы животного к его начальной (в любой предыдущий период) в процентах.
и выражать отношение живой массы животного к его начальной (в любой предыдущий период) в процентах.
![]()
Для более тщательного анализа процесса роста можно сравнивать отношение живой массы в любой последующий период онтогенеза к любому предыдущему:
![]()
Можно также сравнивать общий прирост или прирост в единицу времени (среднесуточный) на одном отрезке онтогенеза к общему (среднесуточному) приросту на любом предыдущем отрезке онтогенеза.
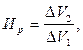 где
где 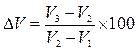
Однако, учитывая неравномерность роста, лучше сравнивать среднесуточный прирост не за один месяц, а за более продолжительный промежуток времени.
Продолжительность роста – показатель способности к росту во времени и определяется возрастом животных, в котором прекращается рост. Для характеристики внутривидовых особенностей роста свиней мы выделяем продолжительность наиболее активного роста, т.е. периода роста, характеризующегося наиболее высокими приростами в единицу времени.
Великорослость – масса (величина) тела, представляющая собой конечный результат роста. При использовании весового метода измерения роста великорослость выражается в килограммах в момент завершения роста животного. Этот критерий можно было бы назвать также «потенцией роста». Однако потенция как наследственно обусловленная возможность роста существует на всем протяжении развития этого процесса.
Скороспелость великорослости – способность выполнять наследственно обусловленную программу роста в наиболее короткий срок. Она применяется при оценке результатов завершенного роста и представляет собой интенсивность роста, выраженную отношением живой массы (общего прироста, прироста в единицу времени) к величине завершенного роста (живая масса взрослых животных, закончивших рост), выраженную в процентах.
![]()
 где
где
![]() - конечная живая масса (величина тела) животных, закончивших рост.
- конечная живая масса (величина тела) животных, закончивших рост.
2. Механизм и ресурсы повышения скорости роста
Механизм и ресурсы повышения скорости роста изучались на основе результатов исследований видовых, породных, половых, индивидуальных особенностей и выявления неизвестных ранее закономерностей роста свиней.
Видовые отличия роста. В отличие от других видов сельскохозяйственных животных, у свиней можно выделить три особенности роста.
Первой особенностью является низкая скорость роста в эмбриональном и высокая в постэмбриональном периоде. По абсолютной скорости роста в эмбриональный период свиньи уступают овцам примерно в 3 раза, крупному рогатому скоту в 12 и лошадям в 15 раз.
В постэмбриональный период по сравнению с эмбриональным скорость роста у свиней увеличивается более, чем в 20 раз, у лошадей – только в 2 раза, а у кроликов – в 5 раз (табл. 1).
1. Особенности роста сельскохозяйственных животных разных видов.
| Вид животных |
Продолжительность роста |
Живая масса, (кг)* |
Среднесуточный прирост |
Кратность увеличения в постэмбриональный период, раз |
|||||
| в эмбриональный период, сут |
в постэмбриональ-ный период, лет |
при рождении |
взрослых животных |
в эмбриональный период |
в постэмбриональ-ный период |
продолжитель-ности роста |
живой массы |
среднесуточного прироста |
|
| Лошади |
340 |
5 |
50 |
500 |
147 |
273 |
5,4 |
10 |
1,8 |
| Крупн. рогат. скот |
280 |
4,5 |
35 |
500 |
125 |
304 |
5,9 |
14,2 |
2,4 |
| Овцы |
150 |
2 |
4 |
40 |
26 |
54 |
4,8 |
10 |
2,0 |
| Свиньи |
114 |
3 |
1,2 |
250 |
10,5 |
228 |
9,6 |
208 |
22,7 |
| Кролики |
30 |
0,8 |
0,06 |
3 |
2 |
10 |
9,7 |
50 |
5 |
| Куры |
21 |
120 сут |
0,04 |
2 |
2 |
17 |
5,7 |
50 |
8,5 |
* Приведена примерная живая масса женских особей
Вторая видовая особенность роста свиней – не сравнимая ни с какими животными
других видов, исключительно высокая интенсивность роста. Интенсивность без преувеличения можно назвать феноменом роста животных этого вида. У свиней она в 15-20 раз выше, чем у самых крупных сельскохозяйственных животных, и в 4 раза выше, чем у кур и кроликов. У лошадей и крупного рогатого скота живая масса к моменту завершения роста увеличивается в сравнении с живой массой при рождении в 10-14 раз, а у свиней – более, чем в 200 раз.
Третьей важной видовой особенностью роста свиней следует считать сочетание высокой интенсивность с большой продолжительностью роста в постэмбриональный период. Так, продолжительность роста в постэмбриональный период больше, чем в эмбриональный у свиней в 9,6 раза, кроликов – 9,7 раза, у лошадей и крупного рогатого скота – 5,3 – 5,9 раза и у овец – в 4,8 раза.
В связи с изложенным следует сделать вывод о целесообразности использования видовых особенностей роста свиней для повышения их скорости роста путем кормления и селекции.
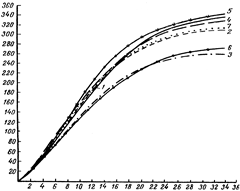
Особенности роста в связи с половым диморфизмом. Процесс роста протекает неодинаково у самцов и самок. Начиная со второго месяца жизни, самцы растут быстрее самок. Наиболее характерной особенностью хряков является их продолжительность активного роста, что особенно заметно на втором и третьем году жизни. Темпы прироста у хряков в возрасте 18-30 месяцев примерно в 2 раза выше, чем у свиноматок. В возрасте, например, 18 месяцев общий прирост хряков примерно на 6 кг превышает месячный прирост свиноматок.
Высокие темпы роста в сочетании с большой продолжительностью активного роста обеспечивают хрякам более высокую живую массу во взрослом состоянии, когда она примерно на 95 кг (38-40%) больше чем у свиноматок. Следовательно, большая продолжительность наиболее активного роста и великорослость – одни из важных биологических резервов повышения скорости роста свиней.
Особенность самок – высокая интенсивность роста в раннем возрасте и низкая в более позднем. Интенсивность роста свинок до 6 месяцев выше, чем у хрячков, после чего интенсивность их роста значительно снижается. Более интенсивный рост самок в раннем возрасте связан с повышенной их скороспелостью и более ранним половым созреванием. Судя по резкому изгибу кривой роста свиноматок в 14-16 месяцев (рис. 1), период резкого ограничения их роста совпадает с возрастом при первом опоросе. У хряков резко выраженное снижение скорости роста наступает в возрасте 20-22 месяцев, что приводит к повышению их великорослости, о чем свидетельствует более высокая живая масса во взрослом состоянии, но снижению их скороспелости по этому признаку. Следовательно, высокая интенсивность роста в раннем возрасте приводит к снижению великорослости и повышению скороспелости свиней.
Породные особенности роста свиней. Свиньи разных пород отличаются по характеру протекания процесса роста и конечным его результатам: скорости, интенсивности, продолжительности активного периода роста, великорослости и скороспелости (табл. 2, рис. 1).
 |
| Вес (кг) |
Возраст (мес.)
Рис. 2. Кривые роста хряков некоторых пород:
1 – крупная белая; 2 – ландрас; 3 – беркширская;
4 – кемеровская; 5 – уржумская; 6 – крупная черная;
7 – короткоухая белая.
2. Изменение веса свиней разных пород с возрастом
| Порода |
Возраст (мес.) |
||||||
| 2 |
4 |
6 |
8 |
10 |
12 |
36 |
|
| вес (кг) |
|||||||
| Хряки |
|||||||
| Крупная белая…………………......... Беркширская……………………….... Брейтовская…………………………. Кемеровская………………………… Белая короткоухая………………….. Крупная черная……………………... Ландрас……………………………… Украинская степная белая…………. Уржумская…………………………... Северокавказская…………………… Сибирская северная………………… Сибирская черно-пестрая породная группа………………………………... |
22 21 20 22 22 20 20 19 20 19 19 19 |
52 49 46 55 54 43 51 43 51 46 46 42 |
83 79 74 92 88 71 86 74 90 76 80 69 |
117 107 103 127 124 99 121 106 130 111 114 99 |
149 135 131 159 160 125 157 138 170 143 146 130 |
181 160 156 189 191 151 188 168 204 172 176 159 |
336 263 268 333 313 273 312 296 347 299 302 304 |
| Свиноматки |
|||||||
| Крупная белая…………………......... Беркширская……………………….... Брейтовская…………………………. Кемеровская………………………… Белая короткоухая………………….. Крупная черная……………………... Ландрас……………………………… Украинская степная белая…………. Уржумская…………………………... Северокавказская…………………… Сибирская северная………………… Сибирская черно-пестрая породная группа………………………………... |
20 18 18 20 20 19 19 18 18 18 17 17 |
48 39 40 51 52 43 46 40 42 39 39 41 |
77 62 66 82 85 70 78 70 76 68 65 71 |
108 86 95 114 117 96 107 99 107 95 93 101 |
137 109 122 144 148 120 133 124 136 120 120 126 |
165 131 146 173 174 142 156 146 162 141 143 147 |
242 180 216 247 244 200 222 223 249 207 225 219 |
Примечание. При определении скорости роста вес поросенка при рождении для всех пород считали равным 1,2 кг.
Скорость роста. Более высокая живая масса отмечается у свиней крупной белой, кемеровской, белой короткоухой, ландрас и уржумской пород. Средняя скорость роста, например хрячков этих пород до 8-месячного возраста колеблется в пределах 479 – 532 г. в сутки. Хрячки других пород в этом возрасте растут со средней скоростью 405 – 467 г. в сутки.
Низкой скоростью роста характеризуются хрячки крупной черной, сибирской черно-пестрой породной группы, брейтовской, украинской степной белой, беркширской пород (среднесуточный прирост 405 – 438 г.).
У хрячков беркширской и крупной черной пород темпы роста достигают максимума на 4-5-м месяце жизни, а у хрячков крупной белой, брейтовской пород и сибирской черно-пестрой породной группы – на 7-8-м месяце. У свинок беркширской, кемеровской, белой короткоухой, крупной черной, ландрас и северокавказской пород кривая темпов роста достигает своего пика на 5-м месяце, у свинок украинской степной белой и уржумской пород – на 6-м, у свинок крупной белой, брейтовской, сибирской северной пород и сибирской черно-пестрой породной группы – на 7-м месяце.
|
|
 Вес (кг)
Вес (кг)
Возраст (мес.)
Рис. 2. Кривые роста хряков некоторых пород:
1 – крупная белая; 2 – ландрас; 3 – беркширская;
4 – кемеровская; 5 – уржумская; 6 – крупная черная;
7 – короткоухая белая.
Таким образом, у свиней беркширской, крупной черной, северокавказский, кемеровской пород период активного роста отмечается в более раннем возрасте, чем у свиней других пород, что свидетельствует об их скороспелости.
Интенсивность роста. По абсолютным показателям прироста трудно судить о напряженности роста. Например, у двух животных, отличающихся по живой массе, одинаковые приросты, скажем за месяц, будут не равнозначны. У свиньи с живой массой 60 кг, давшей общий прирост за месяц, равный 25 кг, интенсивности роста составляет 42%, а у свиньи с живой массой 80 кг и таким же приростом интенсивность роста будет 31%. У животного с низкой живой массой напряженность роста выше, чем у животного с высокой живой массой. Интенсивность роста свиней на примере хряков различных пород, выраженная отношением живой массы в онтогенезе в процентах к живой массе в двухмесячном возрасте, показана в табл. 3.
| Порода |
Отношение живой массы в возрасте 4-12 мес. к живой массе в возрасте 2 мес., % |
||||
| 4 |
6 |
8 |
10 |
12 |
|
| Крупная белая |
237 |
377 |
531 |
677 |
822 |
| Беркширская |
233 |
376 |
509 |
643 |
762 |
| Брейтовская |
230 |
370 |
515 |
655 |
780 |
| Кемеровская |
250 |
418 |
577 |
722 |
859 |
| Белая короткоухая |
245 |
400 |
564 |
727 |
868 |
| Крупная черная |
215 |
355 |
495 |
625 |
755 |
| Ландрас |
255 |
430 |
605 |
785 |
940 |
| Украинская степная белая |
226 |
390 |
557 |
726 |
884 |
| Уржумская |
255 |
450 |
650 |
850 |
1020 |
| Северокавказская |
242 |
400 |
584 |
752 |
905 |
| Сибирская северная |
242 |
421 |
600 |
768 |
926 |
| Сибирская черно-пестрая породн. группа |
221 |
363 |
521 |
684 |
826 |
3. Увеличение живой массы свинок разных пород в онтогенезе по сравнению с живой массой в 2 месяца.
Из таблицы следует, что у скороспелых, но малорослых пород, таких как беркширская, брейтовская, крупная черная, высокая интенсивность роста наблюдается до 6-ти месячного возраста, после чего она начинает снижаться в сравнении с животными многих других пород. У свиней породы ландрас преимущество перед животными других пород по этому признаку роста сохраняется до 10-месячного возраста, а у свиней уржумской породы – до 12-месячного возраста. Свиньи самой многочисленной и распространенной крупной белой породы характеризуются умеренной, но достаточно высокой интенсивностью роста на протяжении всего их периода выращивания.
Продолжительность роста. Анализ кривых роста показывает, что хряки прекращают рост в возрасте 34-35 месяцев, а свиноматки – в возрасте 33-34 месяцев. Свиньи беркширской, брейтовской, крупной черной, северокавказской пород заканчивают рост на 1-2 месяца раньше животных других пород. Примерно на столько же у них короче период наиболее активного роста, который смещен в сторону раннего возраста.
Великорослость. Скорость, интенсивность и продолжительность роста обусловливают ее один важный элемент – великорослость (живая масса животных, закончивших рост). По этому показателю выделяются свиньи крупной белой, уржумской и некоторых других пород. Большой великорослостью обладают свиньи ландрас и короткоухой белой пород. Свиньи украинской степной белой, северокавказской, сибирской северной пород характеризуются средней, а животные беркширской, брейтовской, крупной черной пород – малой великорослостью. Примерно в таком же порядке располагаются породы по скорости и продолжительности периода активного роста.
Скороспелость великорослости. Процесс роста свиней находит выражение в таком важном с хозяйственной точки зрения биологическом свойстве, как скороспелость. Из всех показателей продуктивности и биологических качеств сельскохозяйственных животных скорость роста имеет едва ли не самое непосредственное отношение к скороспелости, ибо как бы ни был широк комплекс биологических особенностей, составляющих это свойство, критерий скорости роста является главнейшим его содержанием. Поэтому не случайно, что во всех или почти всех определениях скороспелости (14, 15, 22 и др), заключен смысл раннего достижения животными высокого уровня производительности без ущерба для их организма.
Скороспелость животных характеризуется ускорением протекания физиологических процессов. Это дает возможность рассматривать некоторые особенности роста свиней как выражение их скороспелости. К таким элементам роста следует отнести скороспелость великорослости, измеряемую отношением показателя роста (например, массы тела) на каком-либо этапе онтогенеза к великорослости как результату роста выраженному в процентах.
![]() где
где
![]() - живая масса животного в искомой точке онтогенеза,
- живая масса животного в искомой точке онтогенеза, ![]() - великорослость как конечный результат роста.
- великорослость как конечный результат роста.
Поскольку у свиней разных пород, а равно как и отдельных индивидов, потенция роста неодинакова, то по массе тела на отдельных этапах развития нельзя судить о том, насколько этот процесс близок к своему завершению. Скороспелость великорослости показывает скорость выполнения наследственно обусловленной программы роста. Результатами сравнения хрячков разных пород по этому показателю приведены в табл. 4.
4. Скороспелость великорослости хрячков разных пород
| Порода |
Возраст (мес.) |
||||||
| 2 |
4 |
6 |
8 |
10 |
12 |
36 |
|
| Отношение веса к весу в возрасте 36 мес. (%) |
|||||||
| Крупная белая…………………......... Беркширская……………………….... Брейтовская…………………………. Кемеровская………………………… Белая короткоухая………………….. Крупная черная……………………... Ландрас……………………………… Украинская степная белая…………. Уржумская…………………………... Северокавказская…………………… Сибирская северная………………… Сибирская черно-пестрая породная группа………………………………... |
6,5 7,9 7,4 6,6 7,0 7,3 6,4 6,4 5,7 6,3 6,6 6,2 |
15,4 18,6 17,1 16,5 17,2 15,7 16,3 14,5 14,7 15,3 15,2 13,8 |
24,7 30,0 27,6 27,6 28,1 26,0 27,5 25,0 25,9 25,4 26,4 22,7 |
34,8 40,6 38,4 38,1 39,6 36,2 38,7 35,7 37,4 37,1 37,7 32,8 |
44,3 51,3 45,1 47,7 51,1 45,7 50,3 46,6 48,7 47,8 48,3 42,7 |
53,8 60,8 58,2 56,7 61,0 55,3 60,2 56,6 58,7 57,5 58,2 52,3 |
100 100 100 100 100 100 100 100 100 100 100 100 |
В раннем возрасте (4-6 мес.) наиболее скороспелы свиньи беркширской, брейтовской, белой короткоухой и крупной черной пород. Высокой скороспелостью отличаются свиньи кемеровской и ландрас пород, о чем свидетельствует высокая относительная масса хрячков в возрасте 6 месяцев. Высокая скороспелость великорослости в 12 месяцев отмечается у хряков беркширской белой короткоухой и ландрас пород.
Скороспелость великорослости свиней крупной белой породы в 12 месяцев ниже, чем у свиней других пород, не считая тугорослых животных сибирской черно-пестрой породной группы. Это свидетельствует о высоком потенциале роста свиней крупной белой породы на втором и третьем годах жизни.
Существует закономерная связь между великорослостью и скороспелостью животных: чем выше интенсивность роста свиней на ранних стадиях онтогенеза и короче фаза активного роста, тем меньше живая масса взрослых животных (великорослость), как общий результат роста. И наоборот, большая великорослость по линии обратной, наследственной связи способствует умеренному и продолжительному росту. Данная взаимозависимость скороспелости и великорослости, очевидно, обусловлена ростом и дифференциацией клеток и выражается в единстве противоречий процесса роста и развития (4, 12, 28), которое можно сформулировать так: дифференциация клеток и тканей организма является следствием ускорения и причиной замедления роста. Следовательно, оценка скорости течения процесса роста животного в связи с конечным его результатом, то есть с величиной, характеризующей законченный рост, может рассматриваться в качестве одного из показателей скороспелости, а великорослость – как важный источник ускорения роста на разных стадиях онтогенеза.
Благодаря такой взаимозависимости и взаимообусловленности, в процессе длительной селекции свиней разных пород сформировались различные модели роста, из которых с учетом классификации (28) соотношений скорости роста и дифференциации клеток можно выделить следующие четыре типа роста свиней:
1) быстрый рост, умеренная скороспелость;
2) быстрый рост, высокая скороспелость;
3) умеренный рост, умеренная скороспелость;
4) умеренный рост, высокая скороспелость.
Животные первого типа характеризуются высокими абсолютными показателями и длительностью активного роста, умеренной скороспелостью и высокой конечной живой массой (великорослость). По такому типу растут и развиваются свиньи крупной белой и уржумской пород.
Особенность второго типа – сравнительно высокие абсолютные и относительные показатели роста. По такому типу растут и развиваются свиньи мясного и беконного направления продуктивности: белой короткоухой, ландрас пород.
Третий тип отличается более низкой в сравнении с первыми двумя типами скоростью роста, что выражается в сравнительно низкой живой массе молодняка, а продолжительный рост обеспечивает животным достаточно высокую живую массу. По такому типу растут свиньи украинской степной белой, сибирской северной и ряд других пород.
Характерную особенность четвертого типа составляют низкие показатели абсолютного и высокие – относительного роста, что выражается в сравнительно низких живой массе и приростах молодых животных, высокой интенсивности роста в раннем возрасте и небольшой конечной живой массе. Наиболее яркие представители такого типа роста свиньи беркширской, брейтовской, крупной черной пород.
В соответствии с указанной классификацией наряду с естественными соотношениями быстрый рост – быстрая дифференциация и медленный рост – медленная дифференциация могут уживаться крайние, противоположные соотношения.
По аналогии с отрицательной корреляцией скорости роста толщины шпика, позволяющей в широкой изменчивости отбирать индивиды с высокой степенью развития обоих признаков и «преодолевать» путем селекции разнонаправленную связь, можно предполагать о трансгрессивной изменчивости соотношений рост – дифференциация.
Наиболее зависимыми, а поэтому широко распространенными соотношениями следует считать те, которые отражают соответствие скорости роста и дифференциации, т.е. второй и третий типы. Обратные соотношения, по-видимому, являются крайними, а следовательно, наиболее редкими вариантами.
Соответствие быстрому росту медленной дифференциации можно объяснить сдвигом в соотношении рост – дифференциация в сторону роста.
Соответствие быстрой дифференциации медленному росту объясняется сдвигом в сторону дифференциации, когда селекция животных на повышенную скороспелость приводит к большой детерминации роста.
Однако следует допускать возможность лишь незначительных сдвигов, ибо в противном случае характер роста мог бы измениться до пределов, превышающих видовые границы. Правда, практика выведения миниатюрных животных (свинья, лошадь, собака) доказывает большие возможности изменения видового роста в сторону уменьшения животных. К сожалению, пока нет примеров столь же значительного изменения видового роста в сторону их увеличения.
Возможность корректирования типов роста свиней доказывает изменение их роста путем селекции в процессе совершенствования пород. Внутрипородные типы роста будут рассмотрены после анализа индивидуальных особенностей роста свиней.
Индивидуальные особенности роста свиней. Количественные изменения роста животного в онтогенезе следует рассматривать в свете действия законов непрерывности, неравномерности и корреляции.
Связь живой массы и привесов в процессе роста. При наличии внутривидового, породного и полового сходства в характере роста свиней существует большая индивидуальная изменчивость (табл. 5).
5. Изменчивость живой массы хряков крупной белой породы в онтогенезе.
| Возраст (мес.) |
М ± m |
|
C ± m |
| 2 4 7 8 9 10 11 12 36 |
21,5 ± 0,44 53,5 ± 0,95 109,0 ± 1,88 126,2 ± 1,69 144,4 ± 1,85 160,1 ± 1,92 177,7 ± 2,14 195,0 ± 1,97 349,2 ± 2,75 |
3,46 ± 0,31 7,48 ± 0,67 14,81 ± 1,33 13,32 ± 1,20 14,60 ± 1,31 15,15 ± 1,36 16,89 ± 1,52 15,52 ± 1,39 21,65 ± 1,94 |
16,14 ± 1,45 13,97 ± 1,25 13,58 ± 1,22 10,55 ± 0,95 10,11 ± 0,91 9,46 ± 0,85 9,50 ± 0,85 7,96 ± 0,71 6,20 ± 0,56 |
Уменьшение коэффициента вариации живой массы с 16,1% в возрасте 2 месяцев до 6,2% в возрасте 36 мес. свидетельствует о большом разнообразии характера роста на ранних стадиях развития. Аналогичная картина наблюдается у женских особей, у которых изменчивость уменьшается с 17,2% в возрасте 4 месяцев до 7,7% в возрасте 36 месяцев.
В целях определения возможностей прогнозирования изменения живой массы свиней на основе предварительной информации целесообразно проанализировать следующие связи: 1) живой массы в отдельные возрастные периоды, 2) живой массы и прироста и 3) прироста на отдельных стадиях онтогенеза. Корреляция живой массы в онтогенезе у хряков крупной белой породы показана в табл. 6.
Связь живой массы сильнее выражена в смежные возрастные периоды и ослабевает по мере удаления коррелирующих признаков друг от друга в процессе роста. Связь усиливается по мере сближения коррелирующих признаков в направлении увеличения возраста животных. Из этого следует, что точность оценки животных по живой массе повышается по мере увеличения их возраста
6. Корреляция живой массы у хряков в онтогенезе*
| Возраст, мес. |
2 |
4 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
36 |
|
|
0,36 |
0,22 |
0,05 |
0,03 |
0,06 |
0,15 |
0,19 |
0,05 |
0,00 |
|
| 4 |
0,64 |
0,60 |
0,38 |
0,37 |
0,34 |
0,30 |
0,41 |
0,26 |
0,00 |
|
| 6 |
0,58 |
0,64 |
0,67 |
0,58 |
0,54 |
0,35 |
0,32 |
0,29 |
0,27 |
|
| 7 |
0,45 |
0,57 |
0,70 |
0,83 |
0,73 |
0,57 |
0,42 |
0,37 |
0,40 |
|
| 8 |
0,42 |
0,61 |
0,65 |
0,81 |
0,89 |
0,72 |
0,46 |
0,50 |
0,38 |
|
| 9 |
0,41 |
0,54 |
0,58 |
0,73 |
0,93 |
0,86 |
0,47 |
0,71 |
0,30 |
|
| 10 |
0,47 |
0,62 |
0,50 |
0,63 |
0,80 |
0,83 |
0,60 |
0,84 |
0,39 |
|
| 11 |
0,37 |
0,48 |
0,49 |
0,54 |
0,69 |
0,69 |
0,73 |
0,56 |
0,22 |
|
|
|
0,35 |
0,50 |
0,52 |
0,56 |
0,77 |
0,79 |
0,80 |
0,77 |
0,35 |
|
| 36 |
0,00 |
0,12 |
0,16 |
0,19 |
0,25 |
0,25 |
0,25 |
0,19 |
0,29 |
 2
2Примечания. 1. По диагонали сверху n = 32, снизу – n = 62. 2. Для числа степеней свободы, равного 30, достоверность корреляции составляет: P < 0,05 при r = 0,35; P < 0,01 при r = 0,45, а равного 60, она составляет P < 0,05 при r = 0,25; P < 0,01 при r = 0,33.
По результатам дисперсионного анализа установлено (табл. 7.), что доля влияния живой массы хряков в возрасте 4 месяцев на их живую массу в 8 месяцев составляет 46%, а в возрасте 36 месяцев она уменьшается до 3%. Доля влияния живой массы в возрасте 10 месяцев на живую массу в 12 и 36 мес. составляет 63 и 19% соответственно.
| Влияющий возраст (мес.) |
Влияемый возраст (мес.) |
|||
| 8 |
10 |
12 |
36 |
|
| влияние организованного фактора (%)
|
||||
| 4 8 10 12 |
46** - - - |
34* 60** - - |
26* 60** 63** - |
3 18 19 23* |
7. Степень влияния живой массы хряков в возрасте 4-12 мес. на их живую массу в последующем ( n = 63)
* P < 0,05.
** P < 0,01.
Примечание. Дисперсионный анализ по методу Н.А. Плохинского (однофакторная схема)
На основании рассмотренных данных можно сделать выводы:
1. Живая масса свиней в процессе их роста находится во взаимосвязи и взаимообусловленности;
2. Сила влияния живой массы в одном периоде онтогенеза животных на их живую массу в другом обратно пропорциональна периоду, в котором она измеряется.
В наших исследованиях не установлена связь между живой массой и среднесуточным приростом в последующие смежные периоды продолжительностью в 1,2 и 3 месяца, что можно объяснить неравномерностью роста животных (табл. 8.).
8. Корреляция среднесуточных приростов молодняка в отдельные возрастные периоды.
| Корреляция привесов по возрастным периодам (мес.) |
Р |
F1 |
||
| свинки (n = 141) |
выращиваемые свинки (n = 135) |
откармливаемые |
||
| свинки (n = 64) |
боровки (n = 55) |
|||
| 1 – 2 и 2 – 4 2 – 4 и 4 – 6 4 – 6 и 6 – 7 1 – 4 и 4 – 7 1 – 4 и 2 – 7 4 – 7 и 2 – 7 |
0,06 ± 0,10 - 0,12 ± 0,10 0,01 ± 0,10 - 0,16 ± 0,10* 0,41 ± 0,09* 0,74 ± 0,05 |
- 0,10 ± 0,08 0,01 ± 0,09 0,02 ± 0,09 - 0,21 ± 0,08 0,39 ± 0,07 0,72 ± 0,04 |
- 0,11 ± 0,13 - 0,11 ± 0,13 0,19 ± 0,21 - 0,02 ± 0,13 0,50 ± 0,10 0,74 ± 0,06 |
- 0,18 ± 0,14 0,02 ± 0,14 - 0,17 ± 0,14 0,22 ± 0,13 0,56 ± 0,10 0,77 ± 0,06 |
* От рождения до 4 месяцев.
Однако высокая и статистически достоверная связь установлена между скоростью роста в каждом из возрастных периодов от 1 до 4 месяцев и от 4 до 7 месяцев, а также в среднем за период выращивания и откорма от 2 до 7 месяцев. Большее влияние на скорость роста за весь период оказал прирост от 4 до 7 месяцев, чем 1 до 4 месяцев, что свидетельствует о влиянии не только продолжительности влияющего периода, но и возраста молодняка.
Вполне очевидно, что скорость роста животных в старшем возрасте по сравнению с ранним имеет большее значение для их общей скорости роста. На основании результатов исследований можно сделать вывод, что генетический прогресс в ускорении роста свиней до 7-месячного возраста может быть обеспечен селекцией на улучшение этого признака как в период в среднем до 4 месяцев, так и от 4 до 7 месяцев.
Неравномерность роста свиней в онтогенезе. Рост свиней в онтогенезе протекает неравномерно. Это выражается в скачкообразности прироста за равные промежутки времени.
Неравномерность роста определяется многими взаимодействующими между собой факторами – условиями внешней среды, изменениями степени процессов ассимиляции и диссимиляции с возрастом животных, разными стадиями и продолжительностью роста органов и тканей, периодичностью накопления клеточного материала и дифференциации клеток, действием и взаимодействием регуляторов роста и т.д.
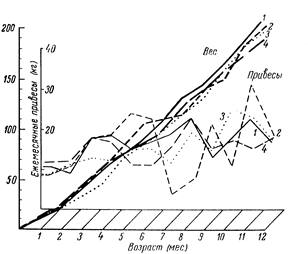 Не затрагивая явлений стадийно-физиологической ритмики, изученной рядом исследователей (13, 25), а имея в виду неравномерность роста, обусловленную неблагоприятными воздействиями окружающей среды (в первую очередь кормлением) можно сказать, что неизменных границ выражения неравномерности роста не существует. Они изменяются в зависимости от силы и продолжительности воздействия влияющих на организм факторов (рис. 3).
Не затрагивая явлений стадийно-физиологической ритмики, изученной рядом исследователей (13, 25), а имея в виду неравномерность роста, обусловленную неблагоприятными воздействиями окружающей среды (в первую очередь кормлением) можно сказать, что неизменных границ выражения неравномерности роста не существует. Они изменяются в зависимости от силы и продолжительности воздействия влияющих на организм факторов (рис. 3).
| Вес (кг) |
Рис. 3. Изменение живой массы и темпов роста у отдельных хряков:
1 – хряк Драчун 2249 (конечный вес 400 кг.); 2 – хряк драчун 8543 (конечный вес 337 кг.); 3 – хряк Кинг 5267 (конечный вес 370 кг.); 4 – хряк Леопард 1459 (конечный вес 300кг.).
Неравномерность проявляется и в росте организма в целом и приросте живой массы. Выражается она в чередовании фаз усиленного и замедленного роста, волнообразном характере ростового процесса с величиной волн разной длины и высоты, влияющих друг на друга и весь результат роста.
Задержка сменяется усилением роста и, наоборот, усиление роста сопровождается замедлением его в последующем.
|
|
 Среднесуточные привесы (г)
Среднесуточные привесы (г)
Возраст (мес.)
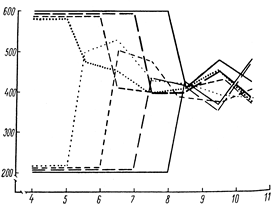
Рис. 4. Схема изменения характера роста свинок в зависимости от темпов роста в предшествующий период.
Закономерности изменения прироста в зависимости от продолжительности ускорения или замедления роста в предшествующем смежном периоде показаны на схеме (4).
Закономерности изменения прироста можно сформулировать так.
1. Чем продолжительнее период высокой скорости роста в предшествующем смежном периоде, тем больше снижение темпов роста в последующем периоде.
2. Чем продолжительнее период замедления роста в предшествующем смежном периоде, тем ниже темпы роста в последующем периоде.
Отличительной особенностью неравномерного роста является взаимосвязь и взаимообусловленность развития этого процесса. Ускорение роста животных тем выше, чем выше степень и короче фаза замедления роста в предшествующем смежном периоде. Но чем выше степень и продолжительность ускорения роста, тем сильнее замедление этого процесса в последующем смежном периоде. Характер течения процесса роста животных на одних стадиях онтогенеза зависит от характера течения его на других.
Чередование фаз ускорения и замедления роста подчиняется следующим закономерностям:
1. Ускорение роста на одной стадии онтогенеза влечет за собой относительное замедление роста на другой.
2. Замедление роста на одной стадии онтогенеза ведет к относительному ускорению роста на другой.
Обратную величину прироста живой массы тела можно рассматривать как проявление механизма регулирования роста, представляющего собой взаимосвязанный и взаимообусловленный процесс увеличения массы тела. Такая направленность и характер прироста выступают в пользу подхода к оценке неравномерности роста организма с позиции регулирующего механизма выполнения генетически обусловленной программы роста.
Изменение режима роста животных приводит к соответствующим изменениям в их организме, направленным на реализацию этой программы. Если рост организма протекает с задержкой во времени, то усиливается действие систем, ускоряющих процессы ассимиляции, вследствие чего нарушенное равновесие в росте восстанавливается. И наоборот, если рост организма перегоняет его хронологический возраст, то вступают в действие сдерживающие ингибирующие системы. Регуляция процесса роста осуществляется путем изменения активности ассимиляции и диссимиляции, в результате чего происходит чередование составляющих его фаз ускорения и замедления.
Увеличение живой массы животных в связи с изменением их прироста на отдельных стадиях онтогенеза осуществляется в соответствии со следующими закономерностями роста и развития:
1. Количественные изменения процесса роста на одной стадии онтогенеза вызывает адекватные изменения на другой.
2. Каждому уровню роста на одной стадии соответствует свой оптимальный уровень на другой.
Внутрипородные типы роста. Под влиянием селекции формируются внутрипородные типы роста свиней, сходные по своей характеристике с породными типами и различающиеся по элементам роста – скорости, интенсивности, великорослости и скороспелости, путем изменения которых можно изменять весь характер течения процесса роста в онтогенезе. Их возникновение обусловлено изменением продолжительности и смещением периода наиболее активного роста в онтогенезе: с повышением интенсивности роста животных в раннем возрасте повышается их скороспелость и уменьшается великорослость, а повышение великорослости приводит к увеличению продолжительности наиболее активного роста и абсолютных показателей скорости. Это становится причиной внутрипородной изменчивости, а также изменения характеристик роста животных в филогенезе (рис. 5,6).
 |
| Вес (кг)
|
Возраст (мес.)
Рис. 5. Изменение кривых роста свиноматок в связи с изменением скорости их роста в отдельные возрастные периоды.
1 – в 2 месяца вес 10 – 11 кг; 2 – в 6 месяцев вес 50 кг; 3 – в 8 месяцев вес 80 кг; 4 – в 2 месяца вес 26 – 27 кг;
5 – в 9 месяцев вес 150 – 159 кг.
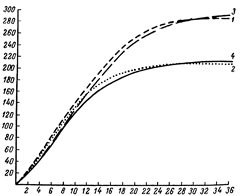
| Живая масса (кг) |
Возраст (мес.)
Рис. 6. Кривые роста свиноматок крупной белой породы с разными типами роста:
1 – быстрый рост, умеренная скороспелость;
2 – быстрый рост, высокая скороспелость;
3 – умеренный рост, умеренная скороспелость;
4 – умеренный рост, высокая скороспелость .
Изучение внутрипородных типов, проведенное на большом поголовье с законченным ростом от 2 до 36 месяцев показали, что каждому типу соответствует свой оптимальный уровень развития процесса роста на каждом этапе онтогенеза. Например, оптимальное изменение живой массы племенных свинок крупной белой породы в разные возрастные периоды находится в следующих границах:
Возраст, мес. Живая масса, кг
2__________________________16 – 25
4__________________________46 – 60
6__________________________60 – 90
7__________________________90 – 119
9__________________________130 – 150
Изменение живой массы животных в более широких пределах, во-первых, не дает желаемых результатов по увеличению скорости роста и, во-вторых, приводит к изменению общих результатов роста. Например, уменьшение живой массы свинок в 2-месячном возрасте менее 15 кг становится причиной задержки роста, некомпенсируемой в течение всей их жизни, а чрезмерное повышение скорости роста в раннем возрасте способствует повышению скороспелости, но снижению великорослости и уровня продуктивности свиноматок.
Изменение процесса роста в онтогенезе у животных, имевших минимальную и максимальную живую массу в возрасте 2 – 9 месяцев, показано в табл. 9. Минимальная и максимальная живая масса в каждом возрастном периоде расположены по диагонали и выделены жирным шрифтом, а по горизонтали показана динамика изменения живой массы у животных данной группы в онтогенезе.
9. Характер роста свинок, имевших минимальную и максимальную живую массу в возрасте 2-9 месяцев.
| Возраст (мес.) |
||||||
| 2 |
4 |
6 |
7 |
8 |
9 |
36 |
| вес (кг) |
||||||
| Рост свинок с минимальной живой массой |
||||||
| 10 – 11 17 16 16 17 18 |
44 30 – 35 44 46 43 42 |
72 60 50 и ниже 58 59 64 |
88 83 79 69 и ниже 70 71 |
99 98 85 81 80 и ниже 77 |
113 115 101 98 99 89 и ниже |
242 252 238 258 260 263 |
| Рост свинок с максимальной живой массой |
||||||
| 26 – 27 23 24 23 22 22 |
53 56 – 60 47 57 55 54 |
88 87 101 – 110 100 96 94 |
101 104 122 120 – 129 120 115 |
115 114 135 136 140 – 149 131 |
127 132 149 153 155 150 – 159 |
251 257 252 258 259 260 |
Примечание. Минимальная и максимальная живая масса в каждом месяце размещены по диагонали.
Данные таблицы наглядно показывают закономерность, заключающуюся в том, что количественные изменения процесса роста на одной стадии онтогенеза вызывают соответствующие изменения на другой. Например, животные, имевшие минимальную живую массу в возрасте 2,4 и 6 месяцев, достигали живой массы 100 кг в возрасте около 9 месяцев, а имевшие минимальную живую массу в период наиболее активного роста свиней крупной белой породы (7-9 месяцев), - за пределами 9 месячного возраста.
Свиньи с максимальной живой массой в 2 и 4 месяца достигли живой массы 100 кг в возрасте 7 месяцев, а в более позднем возрасте (например, 6 месяцев) – достигли живой массы 100 кг в возрасте 6 месяцев.
3. Наследственность и изменчивость признаков роста.
О наследственной обусловленности признаков свидетельствуют достаточно высокие коэффициенты наследуемости живой массы, определенные методом удвоения коэффициентов корреляции между признаками родителей и потомков (табл. 10, 11), и коэффициенты корреляции между живой массой и среднесуточными приростами в отдельные возрастные периоды у сибсов (табл. 12, 13).
Метод определения наследуемости удвоением коэффициента в корреляции основан на промежуточном наследовании признаков и позволяет определять долю разнообразия признака, зависящую от аддитивного действия генов, обусловливающих наследуемость в узком смысле в отличие от наследуемости в широком его значении, включающем другие виды взаимодействия генов (доминирование, эпистаз) в наследовании полигенных признаков.
10. Наследуемость живой массы у свинок на выращивании.
| Наследуемые признаки |
Коэффициенты наследуемости* |
|
| отец-дочь |
мать-дочь |
|
| Живая масса в возрасте 4 мес. |
0,04 |
0,10 |
| Живая масса в возрасте 6 мес. |
0,38 |
0,30 |
| Живая масса в возрасте 7 мес. |
0,48 |
0,36 |
| Живая масса в возрасте 8 мес. |
- |
0,50 |
| Живая масса в возрасте 10 мес. |
- |
0,30 |
| Живая масса в возрасте 12 мес. |
- |
0,46 |
* Определены у 135 пар отец-дочь и 135 пар мать-дочь.
11. Наследуемость живой массы у молодняка на контрольном откорме
| Наследуемые признаки |
Коэффициенты наследуемости |
|||
| у боровков* |
у свинок** |
|||
| отец-сын |
мать-сын |
отец-дочь |
мать-дочь |
|
| Живая масса в 4 мес. |
0,56 |
0,50 |
0,58 |
0,30 |
| Живая масса в 6 мес. |
0,32 |
0,22 |
0,26 |
0,56 |
| Живая масса в 7 мес. |
0,28 |
0,20 |
0,04 |
0,34 |
* Число пар 64.
** Число пар 55.
Надежным доказательством высокой наследуемости признаков роста также служат высокодостоверные коэффициенты корреляции живой массы у свинок – родных сестер крупной белой породы на протяжении всего периода выращивания их до начала племенного использования в два смежных года (табл. 12), а также между среднесуточными приростами у свинок крупной белой породы на выращивании и боровков на откорме в отдельные возрастные периоды (табл. 13).
12. Корреляция живой массы у свинок – сестер.
| Возраст, мес. |
Первый год |
Второй год |
||
| число пар сестер |
коэффициент корреляции |
число пар сестер |
коэффициент корреляции |
|
| 4 |
343 |
0,54 ± 0,06 |
74 |
0,67 ± 0,07 |
| 6 |
321 |
0,63 ± 0,03 |
70 |
0,73 ± 0,05 |
| 7 |
343 |
0,70 ± 0,02 |
68 |
0,68 ± 0,08 |
| 8 |
258 |
0,76 ± 0,02 |
68 |
0,55 ± 0,08 |
| 10 |
258 |
0,70 ± 0,03 |
62 |
0,37 ± 0,11 |
| 12 |
167 |
0,71 ± 0,04 |
45 |
0,59 ± 0,10 |
13. Корреляция среднесуточного прироста в отдельные возрастные периоды у сибсов на выращивании и откорме.
| Продолжительность возрастного периода, мес. |
Свинки-сестры на выращивании |
Боровки-братья на контрольном откорме |
||
| число пар |
коэффициент корреляции |
число пар |
коэффициент корреляции |
|
| 1-2 |
63 |
0,26 ± 0,12 |
24 |
0,05 ± 0,20 |
| 2-4 |
63 |
0,56 ± 0,09 |
24 |
0,15 ± 0,20 |
| 4-6 |
62 |
0,08 ± 0,13 |
24 |
0,34 ± 0,22 |
| 6-7 |
62 |
0,20 ± 0,12 |
20 |
0,11 ± 0,22 |
| 1-4 |
63 |
0,60 ± 0,08 |
24 |
0,34 ± 0,18 |
| 4-7 |
63 |
0,58 ± 0,09 |
22 |
0,31 ± 0,19 |
| 2-7 |
63 |
0,42 ± 0,12 |
22 |
0,18 ± 0,21 |
Высокие коэффициенты корреляции темпов роста у сестер на выращивании отмечались в случаях, когда среднесуточный прирост определяли за возрастные периоды продолжительностью 3-4 месяца и низкие – за более короткие периоды выращивания, что можно объяснить неравномерностью роста животных.
4. Использование признаков роста в селекции в целях повышения скорости роста свиней.
В целях определения результативности использования признаков роста в селекции мы изучали эффективность селекции в зависимости от вида подбора и числа селекционируемых признаков, а также степень влияния отбора и подбора в поколениях.
Как и все количественные признаки, показатели роста подчиняются закономерности аддитивного характера наследования. Эффект селекции по скорости роста (среднесуточный прирост за период выращивания от 2 до 7 месяцев) составил в одном поколении 6,7%, а гетерогенного подбора – 7,5 процента. Селекция на повышение скорости роста способствовала одновременно снижению расхода корма на 1 кг прироста.
Селекцию по скорости роста желательно проводить в сочетании с признаками мясной продуктивности. Преимущества комплексной селекции математически можно выразить так: в случае наличия n
желательных признаков, не связанных между собой, селекция по комплексу признаков будет в![]() раз эффективнее, чем последовательная селекция по одному признаку. Однако комплексная селекция представляется более сложной потому что определяется полигенными влияниями, сложными взаимодействиями генотипа и среды, а тем более, отрицательной корреляцией признаков между собой.
раз эффективнее, чем последовательная селекция по одному признаку. Однако комплексная селекция представляется более сложной потому что определяется полигенными влияниями, сложными взаимодействиями генотипа и среды, а тем более, отрицательной корреляцией признаков между собой.
Результативность селекции изменяется в зависимости от числа селекционируемых признаков, характера и степени влияния их друг на друга (табл. 14).
14. Изменение эффективности отбора крупной белой породы в зависимости от числа селекционируемых признаков ( n = 80).
| Число признаков при отборе |
Признаки и критерии отбора |
Число животных |
Продуктивность |
|||||
| голов |
% ко всей группе |
среднесуточный прирост, г |
толщина шпика на спине, мм |
доля мяса в туше, % |
площадь «мышечного глазка», см2 |
|||
| 1 |
Скорость роста (среднесуточный прирост) 600 г и выше |
42 |
52,5 |
673 |
31 |
58,4 |
29,9 |
|
| 1 |
Толщина шпика на спине 30 мм и менее |
34 |
42,5 |
605 |
27 |
59,9 |
29,6 |
|
| 1 |
Доля мяса в туше 58% и выше |
47 |
58,7 |
594 |
29 |
61,1 |
28,9 |
|
| 1 |
Площадь «мышечного глазка» 30 см2 и более |
21 |
26,2 |
615 |
30 |
61,1 |
32,8 |
|
| 2 |
Скорость роста и толщина шпика |
17 |
21,2 |
659 |
27 |
59,7 |
29,6 |
|
| 3 |
Скорость роста, толщина шпика, доля мяса в туше |
11 |
13,7 |
651 |
27 |
61,9 |
30,3 |
|
| 4 |
Скорость роста, толщина шпика, доля мяса в туше, площадь «мышечного глазка» |
6 |
7,5 |
665 |
27 |
61,0 |
32,8 |
|
По данным таблицы можно сделать выводы:
1. Отбор по одному признаку ведет к улучшению каждого из них;
2. Отбор по скорости роста и признакам мясной продуктивности позволяет поддерживать все признаки отбора на высоком уровне;
3. По мере увеличения числа признаков при отборе резко сокращается число животных, отвечающих требованиям отбора, что свидетельствует о необходимости повышения его жесткости;
4. При отборе по 2, 3 и 4 признакам жесткость отбора животных повышается в 2,5; 3,8 и 7,0 раз в сравнении с отбором только по скорости роста.
Мы изучили также влияние отбора хряков крупной белой породы, а также подбора родительских пар по живой массе на изменение этого признака у их потомков следующих трех поколений. Всего в учет было включено 286 хряков F1 , 97 – F2 , и 45 хряков F3 . Оказалось, что влияние отбора хряков в одном поколении распространяется на потомство хряков следующих двух поколений. Расхождения в живой массе потомков между F1 и F2 составило 5,5%, а между F1 и F3 – 4,7%.
Отбор в двух смежных поколениях обусловил расхождения по указанному признаку в F3 – 4,1% и F4 – 6,6%. Из этого можно сделать вывод, что эффективность отбора повышается по мере увеличения длительности его применения.
Следовательно, обеспечение прогресса в увеличении живой массы хряков возможно только при целенаправленном отборе и подборе на протяжении ряда поколений. Ожидаемый эффект селекции по повышению великорослости хряков на протяжении первых трех поколений может составить 6-7%, а от применения направленного подбора – 7-8%.
5. Разработка методов управления процессом роста и развития в онтогенезе.
5.1. Моделирование процесса роста
Изучение закономерностей роста, источников его ускорения открывает большие возможности для регулирования этого процесса в целях повышения скорости роста, откормочной и мясной продуктивности свиней. С учетом вскрытых резервов ускорения, какими представляются повышение интенсивности, увеличение продолжительности роста, а также великорослости мы спроектировали (1972), а при создании новой скороспелой мясной породы свиней (1998) апробировали оптимальную модель роста, обеспечивающую возможность получения на откорме до 120 кг среднесуточного прироста 850-900 г.
В формировании желательного типа роста как сложного селекционного явления, включающего в себя не один, а целый комплекс признаков, моделирование – наиболее подходящий способ реализации идеи селекции. Правильнее будет сказать, что после создания экстерьерно-типовой модели животного, создание желательного типа роста можно считать вторым прецедентом шаблонной селекции.
Моделирование осуществляется путем комбинирования источников ускорения роста у свиней двух пород с наиболее предпочтительными его типами. Одной из них может быть крупная белая порода (тип роста: быстрый рост, умеренная скороспелость), характеризующаяся высокой скоростью роста до 6-7 – месячного возраста, большой продолжительностью наиболее активного роста (в период примерно от 6 до 12 – месячного возраста) и большой великорослостью. Вторыми могут служить ландрас, белая короткоухая или другие более скороспелые породы (тип роста: быстрый рост, высокая скороспелость), характеризующиеся высокой скоростью роста, скороспелостью великорослости и большой продолжительностью наиболее активного роста. Они более экономичны в сравнении с указанным выше первым типом роста потому, что быстрее растут и менее великорослы.
Модель желательного типа рос та проектируется по принципу совмещения типов роста наиболее прогрессивных пород, характеризующихся большой великорослостью и высокой скороспелостью. Проектируемый тип должен сочетать великорослость одной и высокую скороспелость другой породы.
Моделирование проектируемого типа роста проводится с помощью формулы, предусматривающей использование всех трех источников ускорения этого процесса: интенсивности, длительности активного роста и великорослости сравниваемых пород. Она показывает, как будет изменяться в онтогенезе живая масса одной породы, если она будет нарастать по схеме скороспелости великорослости другой.
Формула имеет следующий вид:
![]() ,
,
где ![]() и
и ![]() - живая масса животных
- живая масса животных ![]() и
и ![]() пород в онтогенезе;
пород в онтогенезе;
![]() и
и ![]() - конечная живая масса (великорослость) животных
- конечная живая масса (великорослость) животных ![]() и
и ![]() пород;
пород;
![]() - показатель скороспелости великорослости породы
- показатель скороспелости великорослости породы ![]() , принятый за 100%.
, принятый за 100%.
Предложим, нам надо узнать, как будет изменяться в онтогенезе живая масса животных породы ![]() , если она будет нарастать по схеме увеличения скороспелости великорослости породы
, если она будет нарастать по схеме увеличения скороспелости великорослости породы ![]() .
.
Пример.
Определить живую массу хрячков породы ![]() в возрасте 6 месяцев, если их скороспелость будет равна скороспелости великорослости хрячков породы
в возрасте 6 месяцев, если их скороспелость будет равна скороспелости великорослости хрячков породы ![]() , у которых живая масса в возрасте 6 месяцев составляет 88 кг и конечная живая масса (великорослость) 313 кг, а конечная живая масса хряков породы
, у которых живая масса в возрасте 6 месяцев составляет 88 кг и конечная живая масса (великорослость) 313 кг, а конечная живая масса хряков породы ![]() будет 336 кг.
будет 336 кг.
Тогда ![]() кг
кг
Если повышение скорости роста животных с проектируемым ростом предусматривается не только путем повышения интенсивности роста (т.е. увеличения скороспелости великорослости), но и путем одновременного увеличения великорослости (конечной живой массы), тогда конечную живую массу породы ![]() (
(![]() ) в уравнении следует заменить на конечную живую массу животных проектируемого типа (
) в уравнении следует заменить на конечную живую массу животных проектируемого типа (![]() ).
).
Тогда формула примет такой вид:
![]()
где ![]() - конечная живая масса животных проектируемого типа.
- конечная живая масса животных проектируемого типа.
5.2. Создание оптимальной модели роста для скороспелой мясной породы СМ-1.
Использованные при моделировании источники ускорения роста (интенсивность, длительность наиболее активного роста и великорослость), разумеется, лучше удается сформировать путем тандемной селекции в одной породе (линии) и полнее использовать в межпородных (линейных) сочетаниях подобно тому, как формируют и применяют отцовские и материнские признаки в системах гибридизации для производства высокопродуктивных товарных гибридов. Нечто подобное происходит и при формировании типов роста, когда накапливаемые в процессе длительной преимущественной селекции признаки ускорения роста удачно сочетаются, нередко усиливая друг друга, в межпородных (линейных) скрещиваниях, быстро являя собой плоды продолжительной селекции. Правильность такой научной концепции, моделирования процесса роста была подтверждена в работе по созданию скороспелой мясной породы (СМ-1). Модель целевого стандарта по признакам роста для создаваемой породы разрабатывали по типам роста наиболее прогрессивных и признанных в мире крупной белой и ландрас пород. Первая из них, характеризуется высокими абсолютными показателями роста, большой великорослостью, продолжительностью активного роста и умеренной скороспелостью. Вторая (ландрас) выделяется высокой скоростью роста и умеренной великорослостью, придающей породе большое совершенство и высокую экономичность.
По нашему замыслу, тип роста крупной белой породы должен был придать создаваемой новой породе большую продолжительность и высокую скорость роста, а породы ландрас – более высокую интенсивность роста в раннем возрасте, хорошую скороспелость, а также экономичность выращивания молодняка и содержания взрослых животных.
Изменение живой массы и скороспелости великорослости животных с наиболее предпочтительными (крупная белая и ландрас породы) и проектируемым (скороспелая мясная порода) типами роста показаны в табл. 15.
| Характеристика роста |
Возраст, мес. |
||||||
| 2 |
4 |
6 |
8 |
10 |
12 |
36 |
|
| Крупная белая порода |
|||||||
| Живая масса, кг |
22 |
52 |
83 |
117 |
149 |
181 |
336 |
| Скороспелость великорослости, %* |
6,5 |
15,4 |
24,7 |
34,8 |
44,3 |
53,8 |
100 |
| Ландрас |
|||||||
| Живая масса, кг |
20 |
51 |
86 |
121 |
157 |
188 |
312 |
| Скороспелость великорослости, % |
6,4 |
16,3 |
27,6 |
38,8 |
50,3 |
60,3 |
100 |
| Проектируемый тип (скороспелая мясная порода) |
|||||||
| Живая масса, кг** |
22 |
55 |
93 |
131 |
170 |
204 |
310 |
| Скороспелость великорослости, % |
7,1 |
17,7 |
30,0 |
42,2 |
54,8 |
65,8 |
100 |
15. Изменение живой массы и скороспелости великорослости у хряков с наиболее предпочтительными и проектируемым типами роста.
Примечания: * Определяли по формуле

* Определяли по формуле
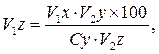 где:
где:
![]() - живая масса животных проектируемого типа (скороспелая мясная) роста в онтогенезе;
- живая масса животных проектируемого типа (скороспелая мясная) роста в онтогенезе;
![]() - живая масса породы ландрас в онтогенезе;
- живая масса породы ландрас в онтогенезе;
![]() - конечная живая масса (великорослость) животных крупной белой породы;
- конечная живая масса (великорослость) животных крупной белой породы;
![]() - скороспелость великорослости животных крупной белой породы принятая за 100%;
- скороспелость великорослости животных крупной белой породы принятая за 100%;
![]() - конечная живая масса (великорослость) животных проектируемого типа роста (скороспелая мясная порода).
- конечная живая масса (великорослость) животных проектируемого типа роста (скороспелая мясная порода).
Таким образом, модель типа роста животных проектируемой скороспелой мясной породы вобрала в себя особенности роста свиней великорослой крупной белой породы и скороспелой породы ландрас. Ее принципиальное отличие от исходных типов заключается в смещении интенсивности роста на более ранний период онтогенеза в сравнении с крупной белой породой и увеличение продолжительности наиболее активного роста на более поздние сроки, чем у свиней породы ландрас. Такое изменение характера протекания процесса роста способствует значительному повышению абсолютной скорости роста у животных проектируемого типа и дает возможность получать при интенсивном откорме желательную реализационную живую массу свиней 120 кг примерно в возрасте 6 месяцев, а 100 кг – в 5,5 месяцев при среднесуточном приросте 800 г и более.
5.3. Практическая реализация теории
Здесь нам остается изложить методы реализации модели роста и показать их эффективность. Их два.
Первый метод это – набор в полигибридных скрещиваниях, сходных по типам роста пород и позволивший сформировать и закрепить в генетическом отношении смоделированный тип роста. Достаточно перечислить исходные породы, принимавшие участие в полигибридных скрещиваниях и послуживших основой для создания новой породы:
- крупная белая (КБ), йоркширская (Й), ландрас (Л), пьетрен (ПТр), миргородская (М), уэссек-сэдлбекская (Ус), эстонская беконная (ЭБ), донской мясной тип (ДМ), белая короткоухая (БК), уэльская (Уэ), крокор (Крок), брейтовская (Бр) и кемеровский мясной тип (КМ). Богатство внесенного в породу генетического вклада будет еще более очевидным, если еще учесть генетическую основу каждой из перечисленных тринадцати лучших отечественных и зарубежных пород, которые в свою очередь, получены на сложной генетической основе. Однако, важно не только количество, но и качество пород, принимавших участие в скрещивании, большое их фенотипическое сходство по уровню и направлению продуктивности, обусловленное тем, что все перечисленные породы создавались и совершенствовались длительное время в мясном направлении.
Наряду с однообразием исходных пород формированию и наследственной консолидации прогрессивного типа роста во многом способствовало применение оригинальных методов отбора, подбора, оценки, селекции и контроля за реализацией селекционной программы (8,9).
Большое значение также имело предусматривавшееся в составе новой породы соотношение долей крови участвовавших в скрещивании пород: 25% крови крупной белой породы и 75% ландрас и других, сходных с ней, пород.
Второй способ воплощения научной идеи по ускорению роста это – включение основных критериев роста в целевой стандарт породы, служивший основой для применения метода селекции по независимым уровням – фенотипическому сходству. Стандартом предусматривалось, во-первых, повышение скорости примерно до 800 г в сутки и, во-вторых, проводить селекцию не до 100 кг, как это принять в отечественном свиноводстве, а до 120 кг, увеличив продолжительность откорма в зоне наиболее активного роста на 20-25 суток, что служит источником ускорения роста.
Результаты селекции в процессе создания новой породы представлены в табл. 16.
16. Изменение скорости роста, откормочной и мясной продуктивности свиней скороспелой мясной породы (СМ-1) в процессе ее создания.
| Селекционный признак |
Исходные формы |
Скороспелая мясная порода в поколениях |
Селекционный сдвиг |
Эффект селекции за одно поколение |
|||
| F1 |
F2 |
F3 |
F4 |
||||
| Возраст в 120 кг, сут. |
209 |
212 |
226 |
194 |
188 |
21 |
5,25 |
| Среднесуточный прирост, г |
767 |
733 |
742 |
837 |
847 |
80 |
20 |
| Расход корма на 1 кг прироста, корм. ед. |
3,79 |
3,46 |
3,51 |
3,27 |
3,38 |
0,41 |
0,10 |
| Выход мяса в туше, % |
56,9 |
54,6 |
56,1 |
57,7 |
58,0 |
1,1 |
0,27 |
Достигнуты высокие результаты в селекции по ускорению роста и улучшению связанных с ним признаков, создана новая скороспелая мясная порода свиней, соответствующая по продуктивности уровню мировых селекционных достижений: свиньи новой породы на интенсивном откорме достигают живой массы 120 кг в возрасте 188 суток, дают среднесуточный прирост 847 г и туши с выходом постного мяса 58%, при затратах 3,38 корм. ед. на 1 кг прироста.
Свиньи из племхозов «Дружба» Ленинградской области и «Петровский» Ставропольского края дали среднесуточный прирост по 948 г и 953 г соответственно, израсходовав по 3,27 и 3,25 корм. ед. на 1 кг прироста, и туши (при убое животных в 120 кг) с выходом постного мяса по 55,4 процента.
Результаты сравнительной оценки скорости роста, откормочной и мясной продуктивности свиней скороспелой мясной (СМ-1), крупной белой и ландрас пород представлены в табл. 17.
17. Скорость роста, откормочная и мясная продуктивность
свиней скороспелой мясной, крупной белой и ландрас пород.
| Порода |
Возраст в 120 кг, сут. |
Среднесуточный прирост, г |
Расход корма на 1 кг прироста, корм. ед. |
Содержится в туше, % |
|
| мяса |
сала |
||||
| Скороспелая мясная (СМ-1) |
188 |
847 |
3,38 |
58,0 |
22,1 |
| Крупная белая |
221 |
713 |
3,70 |
55,0 |
26,3 |
| Ландрас |
215 |
730 |
3,62 |
57,6 |
24,9 |
Сравнительные испытания показали, что по скорости роста и связанным с ней признакам откормочной и мясной продуктивности свиньи скороспелой мясной породы (СМ-1) значительно превосходят животных многих отечественных и зарубежных, в том числе крупной белой и ландрас. При откорме до 120 кг их продуктивность была выше, чем свиней крупной белой породы и ландрас по величине среднесуточного прироста на 134 и 117 г, расходу корма на 1 кг прироста – на 0,32 и 0,24 корм. ед. и содержанию мяса в туше – на 3 и 0,4% соответственно. Все это дает основание считать селекцию по признакам роста эффективной.
Заключение
Теория высокой скорости роста была положена в основу создания скороспелой мясной породы свиней, соответствующей уровню лучших мировых селекционных достижений (8). Практическая реализация селекционной программы по выведению новой породы с высокой скоростью роста осуществлялась двумя методами:
1. путем выбора исходных пород с желательным типом роста и
2. увеличения продолжительности периода наиболее активного роста путем повышения реализационной живой массы свиней до 120 кг в процессе целенаправленной селекции.
Из 13-ти принимавших участие в полигибридных скрещиваниях лучших отечественных и зарубежных пород генетический вклад пород с быстрым ростом и высокой скороспелостью в общий генофонд новой породы составлял примерно 77% и пород с типом быстрый рост, умеренная скороспелость – 23%.
Повышение реализационной живой массы свиней до 120 кг позволило увеличить продолжительность наиболее активного роста и повысить скорость роста свиней на интенсивном откорме до 800-900 г в сутки.
Библиографический список
1. Балинский Б.И. Рост и формообразование Сб. «Рост животных». Биомедгиз, М., 1935.
2. Борисенко Е.Я., Евстратова А.М. О типах роста животных. Известия ТСХА, №4, 1965.
3. Вермель Е.М. Величина и рост клеток. Сб. «Рост животных», Биомедгиз, М., 1935.
4. Гофман-Кадошников П.Б. Основные итоги изучения в СССР закономерностей роста организма за тридцать лет. «Успехи современной биологии», 1947, т. XXIV, в 3 (6).
5. Жинкин Л.Н. Дифференциация и продолжительность жизни клеток. Руководство по цитологии. М. – Л., 1966, т. II.
6. Кабанов В.Д. Рост и мясные качества свиней. Изд. «Колос», М., 1972, 191 с.
7. Кабанов В.Д. Биологические основы повышения скорости роста и улучшения мясных качеств в свиней. Диссертация на соискание ученой степени доктора сельскохозяйственных наук. М., 1973, 352 с.
8. Кабанов В.Д., Гупалов Н.В., Епишин В.А., Кошель П.П. Теория и методы выведения скороспелой мясной породы свиней. Изд. ВНИИплем, М., 1998, 380 с.
9. Кабанов В.Д. Гомогенизация наследственности путем дробления степени кровности и ее роль в селекции сельскохозяйственных животных. М.: МГАВМиБ им. К.И. Скрябина, 2001, 27 с.
10. Комфорт А. Биология старения. – М. – «Мир», 1967.
11. Калугин Н.В. О ритмичности роста поросят. Тр. Оренбургского СХИ, Оренбург, 1966, т.19.
12. Колесник Н.Н. Измерение роста и скороспелости животных. Тр. Института генетики АН СССР, 1939, № 13.
13. Курбатов А.И. Показатели скороспелости свиней и факторы их обусловливающие. «Известия Иркутского СХИ», Иркутск, 1969, т. 1, вып. 27.
14. Ладан П.Е., Белкина Н.Н. Новый метод определения скороспелости свиней. Ж. «Свиноводство», 1961, № 7.
15. Викторов П.И. Скороспелость с-х животных и пути ее повышения. Краснодар, 1966.
16. Ларк К.Г. Регуляция биосинтеза ДНК в клетке. В кн.: «Молекулярная генетика», М., 1964.
17. Майнот Ч.С. Современные проблемы биологии. М., 1965
18. Малигонов А.А. О скорости весового роста животного организма в различные периоды в связи с величиной растущей массы. Тр. «Кубанского СХИ», 1926, т. III.
19. Мильман М.С. Учение о росте, старении и смерти. Баку, 1926.
20. Никитин В.Н. О некоторых основных факторах онтогенеза. «Ведущие проблемы возрастной физиологии и биохимии», М., 1966.
21. Плохинский Н.А. Наследуемость. Новосибирск. Изд-во СО АН СССР, 1964.
22. Свечин К.Б. Индивидуальное различие сельскохозяйственных животных. Изд. АН УССР, Киев, 1961.
23. Сипачев С.Г. Ритмичность роста животных. Тюмень, 1970.
24. Уоддингтон К. Морфогенез и генетика. М., 1964.
25. Федоров В.И. Ритмичность роста и ее практическое значение. Ж. «Животноводство», 1958, № 3.
26. Хэммонд Дж. Биологические проблемы животноводства. М., «Колос», 1964.
27. Чирвинский Н.П. Изменение сельскохозяйственных животных под влиянием обильного и скудного кормления в молодом возрасте. Избранные сочинения, 1949, т.1.
28. Шмальгаузен И.И. Определение основных понятий и методика исследования роста. Сб. «Рост животных», Биомедгиз. 1935.
29. Якушев В.И. О ритмичном характере роста свиней. Тр. Всесоюзн. н.-и. экспериментальной ветеринарии. 1957. т. 20.