| Похожие рефераты | Скачать .docx | Скачать .pdf |
Дипломная работа: Структурно-функциональная организация генетического материала
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА
1. Наследственность и изменчивость - фундаментальные свойства живого
2. История формирования представлений об организации материального субстрата наследственности и изменчивости
3. Общие свойства генетического материала и уровни организации генетического аппарата
4. Генный уровень организации генетического аппарата
4.1 Химическая организация гена
4.1.1 Структура ДНК. Модель Дж. Уотсона и Ф. Крика
4.1.2 Способ записи генетической информации в молекуле ДНК. Биологический код и его свойства
4.2 Свойства ДНК как вещества наследственности и изменчивости
4.2.1 Самовоспроизведение наследственного материала. Репликация ДНК
4.2.2 Механизмы сохранения нуклеогидной последовательности ДНК. Химическая стабильность. Репликация. Репарация
4.2.3 Изменения нуклеотидных последовательностей ДНК.
4.2.4 Элементарные единицы изменчивости генетического материала. Мутон. Рекон
4.2.5 Функциональная классификация генных мутаций
4.2.6 Механизмы, снижающие неблагоприятный эффект генных мутаций
4.3 Использование генетической информации в процессах жизнедеятельности
4.3.1 Роль РНК в реализации наследственной информации
4.3.2 Особенности организации и экспрессии генетической информации у про - и эукариот
1. Наследственность и изменчивость - фундаментальные свойства живого
Жизнь как особое явление характеризуется продолжительностью существования во времени (на Земле она возникла более 3,5 млрд. лет назад), что обеспечивается преемственностью поколений живых систем. Происходит смена поколений клеток в организме, смена поколений организмов в популяциях, смена видов в системе биоценоза, смена биоценозов, образующих биосферу. В основе непрерывного существования жизни во времени лежит способность живых систем к самовоспроизведению. Сохранение жизни в меняющихся условиях оказывается возможным благодаря эволюции живых форм, в процессе которой у них появляются изменения, обеспечивающие приспособление к новой среде обитания. Непрерывность существования и историческое развитие живой природы обусловлены двумя фундаментальными свойствами жизни: наследственностью и изменчивостью.
В учебных курсах свойства наследственности и изменчивости традиционно рассматривают относительно клетки и организма. На самом деле они проявляются и на надорганизменных уровнях. На клеточном и организменном (онтогенетическом) уровнях организации живого под наследственностью понимают свойство клеток или организмов в процессе самовоспроизведения передавать новому поколению способность к определенному типу обмена веществ и индивидуального развития, в ходе которого у них формируются общие признаки и свойства данного типа клеток и вида организмов, а также некоторые индивидуальные особенности родителей. На популяционно-видовом уровне организации жизни наследственность проявляется в поддержании постоянного соотношения различных генетических форм в ряду поколений организмов данной популяции (вида). На биоценотическом уровне продолжительное существование биоценоза обеспечивается сохранением определенных соотношений видов организмов, образующих этот биоценоз.
В ходе возникновения и развития жизни на Земле наследственность играла решающую роль, так как закрепляла в ряду поколений биологически полезные эволюционные приобретения, обеспечивая определенный консерватизм организации живых систем. Наследственность является одним из главных факторов эволюции.
Продолжительное существование живой природы во времени на фоне меняющихся условий было бы невозможным, если бы живые системы не обладали способностью к приобретению и сохранению некоторых изменений, полезных в новых условиях среды. Свойство живых систем приобретать изменения и существовать в различных вариантах называется изменчивостью.
У отдельных клеток и организмов одного вида изменчивость, затрагивая их индивидуальное развитие, проявляется в возникновении отличий между ними. На популяционно-видовом уровне организации жизни это свойство проявляется в наличии генетических различий между отдельными популяциями вида, что лежит в основе образования новых видов. Появление новых видов вносит изменения в межвидовые взаимоотношения в биоценозах. Изменчивость в определенном смысле отражает динамичность организации живых систем и наряду с наследственностью является ведущим фактором эволюции. Несмотря на то что по своим результатам наследственность и изменчивость разнонаправлены, в живой природе эти два фундаментальных свойства образуют неразрывное единство, чем достигается одновременно сохранение в процессе эволюции имеющихся биологически целесообразных качеств и возникновение новых, делающих возможным существование жизни в разнообразных условиях.
2. История формирования представлений об организации материального субстрата наследственности и изменчивости
Наследственность и изменчивость как важнейшие свойства любой живой системы обеспечиваются функционированием особого материального субстрата. В ходе исторического развития биологической науки представления о его свойствах, организации и химической природе постоянно расширяются и усложняются.
В 60-х гг. XIX в. основоположник генетики (науки о наследственности и изменчивости) Г. Мендель (1865) высказал первые предположения об организации наследственного материала. На основании результатов своих экспериментов на горохе он пришел к заключению, что наследственный материал дискретен, т.е. представлен отдельными наследственными задатками, отвечающими за развитие определенных признаков организмов. По утверждению Менделя, в наследственном материале организмов, размножающихся половым путем, развитие отдельного признака обеспечивается парой аллельных задатков, пришедших с половыми клетками от обоих родителей. При образовании гамет в каждую из них попадает лишь один из пары аллельных задатков, поэтому гаметы всегда "чисты". В 1909 г.В. Иогансен назвал "наследственные задатки" Менделя генами.
80-е гг. XIX в. ознаменовались важными достижениями в области цитологии: были описаны митоз и мейоз - деление соответственно соматических и половых клеток, в ходе которых закономерно между дочерними клетками распределяются ядерные структуры - хромосомы (В. Вольдейер, 1888).
Данные о характере распределения хромосом в процессе клеточного деления позволили в начале XX в.Т. Бовери (1902-1907) и У. Сетгону (1902-1903) сделать вывод о том, что преемственность свойств в ряду поколений клеток и организмов определяется преемственностью их хромосом. Хромосомы стали рассматривать как материальные носители наследственной программы.
Дальнейшая разработка хромосомной теории наследственности, объединяющей представления о наследственных задатках и хромосомах, была осуществлена в начале XX в. Т. Морганом и его сотрудниками. В опытах, выполненных на дрозофиле, было подтверждено ранее высказанное предположение о роли хромосом в обеспечении наследственности. Установлено, что гены размещаются в хромосомах, располагаясь в них в линейном порядке. Гены каждой хромосомы образуют группу сцепления, число которых определяется количеством хромосом в половых клетках. Гены одной группы сцепления наследуются, как правило, совместно. Однако в ряде случаев происходит их перекомбинация в связи с кроссинговером, частота которого зависит от расстояния между генами.
Таким образом, в хромосомной теории нашел отражение один из важнейших принципов генетики - единство дискретности и непрерывности наследственного материала.
Необходимо отметить, что также в начале XX в. были обнаружены факты, которые доказывали наличие в клетках внехромосомного наследственного материала, располагающегося в различных цитоплазматических структурах и определяющего особую цитоплазматическую наследственность (К. Корренс, 1908).
Примерно в это же время X. де Фризом (1901) были заложены основы учения о мутационной изменчивости, связанной с внезапно возникающими изменениями в наследственных задатках или хромосомах, что приводит к изменениям тех или иных признаков организма. В последующие годы было обнаружено мутагенное действие на хромосомы и гены рентгеновских лучей, радиационного излучения, определенных химических веществ и биологических агентов.
В результате этих исследований стало очевидным, что наследственность и изменчивость обусловлены функционированием одного и того же материального субстрата.
В первые десятилетия XX в. были получены данные, свидетельствующие в пользу зависимости состояния признаков от характера взаимодействия генов, что выходило за рамки отношений доминантности и рецессивности, описанных еще Менделем. Отсюда появилось представление о генетическом аппарате как о системе взаимодействующих генов - генотипе, который сосредоточен в хромосомном наборе - кариотипе.
Изучение химического состава хромосом выявило два основных вида соединений, образующих эти структуры, - белки и нуклеиновые кислоты. В первой половине XX в. исследователями решался вопрос о химической природе субстрата наследственности и изменчивости. Первоначально высказывались предположения в пользу белков. В 1928 г.Ф. Гриффитом был поставлен опыт на пневмококках, в котором наблюдалось изменение (трансформация) некоторых наследственных свойств одного бактериального штамма под влиянием материала, полученного из убитых клеток другого штамма. Химическая природа вещества, трансформирующего наследственные свойства бактерий, была установлена лишь в 1944 г.О. Эйвери, доказавшим его принадлежность к нуклеиновым кислотам (ДНК).
Другими доказательствами участия ДНК в обеспечении наследственности и изменчивости являются:
1) постоянство содержания ДНК во всех типах соматических клеток организма;
2) соответствие содержания ДНК плоидности клеток (в соматических клетках ее вдвое больше, чем в половых, в полиплоидных клетках оно соответствует количеству наборов хромосом);
3) явление генетической рекомбинации у бактерий при их конъюгации, в ходе которой осуществляется проникновение части ДНК из одной клетки в другую и изменение свойств последней;
4) изменение наследственных свойств бактериальных клеток путем переноса ДНК от одного штамма к другому с помощью ДНК-фага - явление трансдукции;
5) инфицирующая активность изолированной нуклеиновой кислоты вирусов.
Важным результатом целенаправленного изучения нуклеиновых кислот было создание Дж. Уотсоном и Ф. Криком (1953) пространственной модели молекулы ДНК.
Во второй половине XX в. усилия ученых направлены на изучение свойств нуклеиновых кислот, составляющих основу их генетических функций, способов записи и считывания наследственной информации, характера и структуры генетического кода, механизмов регуляции активности генов в процессе формирования отдельных признаков и фенотипа в целом. В 60-х гг. работами М. Ниренберга, С. Очоа, X. Кораны и других была произведена полная расшифровка генетического кода, установлено соответствие триплетов нуклеотидов в молекуле нуклеиновых кислот определенным аминокислотам. В 70-х гг. стали активно разрабатываться методы генной инженерии, позволяющие целенаправленно изменять наследственные свойства живых организмов.
К концу XX столетия, благодаря новым молекулярно-генетическим технологиям, появилась возможность определять последовательности нуклеотидов в молекулах ДНК геномов различных организмов (прочтение ДНК-текстов). ДНК-тексты генома человека, представленные в целом 3 млрд. пар нуклеотидов, в основном прочитаны к 2001 году. Научно-практическое направление молекулярной биологии, имеющее целью определение нуклеотидных последовательностей молекул ДНК, получило название геномики.
3. Общие свойства генетического материала и уровни организации генетического аппарата
На основании приведенных выше определений наследственности и изменчивости можно предположить, каким требованиям должен отвечать материальный субстрат этих двух свойств жизни.
Во-первых, генетический материал должен обладать способностью к самовоспроизведению, чтобы в. процессе размножения передавать наследственную информацию, на основе которой будет осуществляться формирование нового поколения. Во-вторых, для обеспечения устойчивости характеристик в ряду поколений наследственный материал должен сохранять постоянной свою организацию. В-третьих, материал наследственности и изменчивости должен обладать способностью приобретать изменения и воспроизводить их, обеспечивая возможность исторического развития живой материи в меняющихся условиях. Только в случае соответствия указанным требованиям материальный субстрат наследственности и изменчивости может обеспечить длительность и непрерывность существования живой природы и ее эволюцию.
Современные представления о природе генетического аппарата позволяют выделить три уровня его организации: генный, хромосомный и геномный. На каждом из них проявляются основные свойства материала наследственности и изменчивости и определенные закономерности его передачи и функционирования.
4. Генный уровень организации генетического аппарата
Элементарной функциональной единицей генетического аппарата, определяющей возможность развития отдельного признака клетки или организма данного вида, является ген (наследственный задаток, по Г. Менделю). Передачей генов в ряду поколений клеток или организмов достигается материальная преемственность - наследование потомками признаков родителей.
Под признаком понимают единицу морфологической, физиологической, биохимической, иммунологической, клинической и любой другой дискретности организмов (клеток), т.е. отдельное качество или свойство, по которому они отличаются друг от друга.
Большинство перечисленных выше особенностей организмов или клеток относится к категории сложных признаков, формирование которых требует синтеза многих веществ, в первую очередь белков со специфическими свойствами - ферментов, иммунопротеинов, структурных, сократительных, транспортных и других белков. Свойства белковой молекулы определяются аминокислотной последовательностью ее полипептидной цепи, которая прямо задается последовательностью нуклеотидов в ДНК соответствующего гена и является элементарным, или простым, признаком.
Основные свойства гена как функциональной единицы генетического аппарата определяются его химической организацией,
4.1 Химическая организация гена
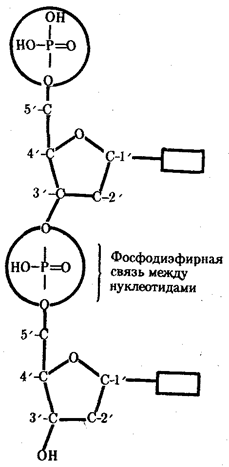
Исследования, направленные на выяснение химической природы наследственного материала, неопровержимо доказали, что материальным субстратом наследственности и изменчивости являются нуклеиновые кислоты, которые были обнаружены Ф. Мишером (1868) в ядрах клеток гноя. Нуклеиновые кислоты являются макромолекулами, т.е. отличаются большой молекулярной массой. Это полимеры, состоящие из мономеров - нуклеотидов, включающих три компонента: сахар (пентозу), фосфат и азотистое основание (пурин или пиримидин). К первому атому углерода в молекуле пентозы С-1' присоединяется азотистое основание (аденин, гуанин, цитозин, тимин или урацил), а к пятому атому углерода С-5' с помощью эфирной связи - фосфат; у третьего атома углерода С-3' всегда имеется гидроксильная группа - ОН (рис.1).
Соединение нуклеотидов в макромолекулу нуклеиновой кислоты происходит путем взаимодействия фосфата одного нуклеотида с гидроксилом другого так, что между ними устанавливается фосфодиэфирная связь (рис.2). В результате образуется полинуклеотидная цепь. Остов цепи состоит из чередующихся молекул фосфата и сахара. К молекулам пентозы в положении С-1' присоединено одно из перечисленных выше азотистых оснований (рис.3).

Рис.1. Схема строения нуклеотида
Объяснение см. в тексте; обозначения компонентов нуклеотида, использованные в этом рисунке, сохраняются во всех последующих схемах нуклеиновых кислот
Сборка полинуклеотидной цепи осуществляется при участии фермента полимеразы, который обеспечивает присоединение фосфатной группы следующего нуклеотида к гидроксильной группе, стоящей в положении 3', предыдущего нуклеотида (рис. 3.3). Благодаря отмеченной специфике действия названного фермента наращивание полинуклеотидной цепи происходит только на одном конце: там, где находится свободный гидроксил в положении 3'. Начало цепи всегда несет фосфатную группу в положении 5'. Это позволяет выделить в ней 5' и 3 '-концы.
Среди нуклеиновых кислот различают два вида соединений: дезоксирибонуклеиновую (ДНК) и рибонуклеиновую (РНК) кислоты. Изучение состава основных носителей наследственного материала - хромосом - обнаружило, что их наиболее химически устойчивым компонентом является ДНК, которая представляет собой субстрат наследственности и изменчивости.
Рис.2. Схема соединения нуклеотидов в полинуклеотидную цепь |
Рис.3. Схема полинуклеотидной цепи (стрелкой указано направление роста цепи) Присоединение очередного нуклеотида сопровождается образованием эфирной связи между ОН-группой на 3'-конце полинуклеотидной цепи и фосфатом свободного нуклеотида, остальные пояснения см. в тексте |

4.1.1 Структура ДНК. Модель Дж. Уотсона и Ф. Крика
ДНК состоит из нуклеотидов, в состав которых входят сахар - дезоксирибоза, фосфат и одно из азотистых оснований - пурин (аденин или гуанин) либо пиримидин (тимин или цитозин). Особенностью структурной организации ДНК является то, что ее молекулы включают две полинуклеотидные цепи, связанные между собой определенным образом. В соответствии с трехмерной моделью ДНК, предложенной в 1953 г. американским биофизиком Дж. Уотсоном и английским биофизиком и генетиком Ф. Криком, эти цепи соединяются друг с другом водородными связями между их азотистыми основаниями по принципу комплементарности. Аденин одной цепи соединяется двумя водородными связями с тимином другой цепи, а между гуанином и цитозином разных цепей образуются три водородные связи. Такое соединение азотистых оснований обеспечивает прочную связь двух цепей и сохранение равного расстояния между ними на всем протяжении.

Рис.4. Схема строения молекулы ДНК. Стрелками обозначена антилараллельность целей.
Другой важной особенностью объединения двух полинуклеотидных цепей в молекуле ДНК является их антипараллельность: 5'-конец одной цепи соединяется с 3'-концом другой, и наоборот (рис.4).
Данные рентгеноструктурного анализа показали, что молекула ДНК, состоящая из двух цепей, образует спираль, закрученную вокруг собственной оси. Диаметр спирали составляет 2 нм, длина шага - 3, 4 нм. В каждый виток входит 10 пар нуклеотидов.
Чаще всего двойные спирали являются правозакрученными - при движении вверх вдоль оси спирали цепи поворачиваются вправо. Большинство молекул ДНК в растворе находится в правозакрученной - В-форме (В-ДНК). Однако встречаются также левозакрученные формы (Z-ДНК). Какое количество этой ДНК присутствует в клетках и каково ее биологическое значение, пока не установлено (рис.3.5).

Рис.5. Пространственные модели левозакрученной Z-формы (I) и правозакрученной В-формы (II) ДНК
Таким образом, в структурной организации молекулы ДНК можно выделить первичную структуру - полинуклеотидную цепь, вторичную структуру-две комплементарные друг другу и антипараллельные полинуклеотидные цепи, соединенные водородными связями, и третичную структуру - трехмерную спираль с приведенными выше пространственными характеристиками.
4.1.2 Способ записи генетической информации в молекуле ДНК. Биологический код и его свойства
Первично все многообразие жизни обусловливается разнообразием белковых молекул, выполняющих в клетках различные биологические функции. Структура белков определяется набором и порядком расположения аминокислот в их пептидных цепях. Именно эта последовательность аминокислот в пептидах зашифрована в молекулах ДНК с помощью биологического (генетического) кода. Относительная примитивность структуры ДНК, представляющей чередование всего лишь четырех различных нуклеотидов, долгое время мешала исследователям рассматривать это соединение как материальный субстрат наследственности и изменчивости, в котором должна быть зашифрована чрезвычайно разнообразная информация.
В 1954 г.Г. Гамовым было высказано предположение, что кодирование информации в молекулах ДНК должно осуществляться сочетаниями нескольких нуклеотидов. В многообразии белков, существующих в природе, было обнаружено около 20 различных аминокислот. Для шифровки такого их числа достаточное количество сочетаний нуклеотидов может обеспечить лишь триплетный код, в котором каждая аминокислота шифруется тремя стоящими рядом нуклеотидами. В этом случае из четырех нуклеотидов образуется 43 = 64 триплета. Код, состоящий из двух нуклеотидов, дал бы возможность зашифровать только 42 = 16 различных аминокислот.
Полная расшифовка генетического кода проведена в 60-х гг. нашего столетия. Из 64 возможных триплетов ДНК 61 кодирует различные аминокислоты; оставшиеся 3 получили название бессмысленных, или "нонсенс-триплетов". Они не шифруют аминокислот и выполняют функцию знаков препинания при считывании наследственной информации. К ним относятся АТТ, АЦТ, АТЦ.
Обращает на себя внимание явная избыточность кода, проявляющаяся в том, что многие аминокислоты шифруются несколькими триплетами (Рис.6). Это свойство триплетного кода, названное вырожденностью, имеет очень важное значение, так как возникновение в структуре молекулы ДНК изменений по типу замены одного нуклеотида в полинуклеотидной цепи может не изменить смысла триплета. Возникшее таким образом новое сочетание из трех нуклеотидов кодирует ту же самую аминокислоту.
В процессе изучения свойств генетического кода была обнаружена его специфичность. Каждый триплет способен кодировать только одну определенную аминокислоту. Интересным фактом является полное соответствие кода у различных видов живых организмов. Такая универсальность генетического кода свидетельствует о единстве происхождения всего многообразия живых форм на Земле в процессе биологической эволюции. Незначительные отличия генетического кода обнаружены в ДНК митохондрий некоторых видов. Это не противоречит в целом положению об универсальности кода, но свидетельствует в пользу определенной дивергентности в его эволюции на ранних этапах существования жизни. Расшифровка кода в ДНК митохондрий различных видов показала, что во всех случаях в митохондриальных ДНК отмечается общая особенность: триплет АЦТ читается как АЦЦ, и поэтому из нонсенс-триплета превращается в шифр аминокислоты триптофана.

Рис.6. Аминокислоты и кодирующие их триплеты ДНК
Другие особенности являются специфичными для различных видов организмов. У дрожжей триплет ГАТ и, возможно, все семейство ГА кодирует вместо аминокислоты лейцина треонин. У млекопитающих триплет ТАГ имеет то же значение, что и ТАЦ, и кодирует аминокислоту метионин вместо изолейцина. Триплеты ТЦГ и ТЦЦ в ДНК митохондрий некоторых видов не кодируют аминокислот, являясь нонсенс-триплетами. Наряду с триплетностью, вырожденностью, специфичностью и универсальностью важнейшими характеристиками генетического кода являются его непрерывность и неперекрываемость кодонов при считывании. Это означает, что последовательность нуклеотидов считывается триплет за триплетом без пропусков, при этом соседние триплеты не перекрывают друг друга, т.е. каждый отдельный нуклеотид входит в состав только одного триплета при заданной рамке считывания (рис.3.7). Доказательством неперекрываемости генетического кода является замена только одной аминокислоты в пептиде при замене одного нуклеотида в ДНК. В случае включения нуклеотида в несколько перекрывающихся триплетов его замена влекла бы за собой замену 2-3 аминокислот в пептидной цепи.

Рис.7. Непрерывность и непререкаемость генетического кода при считывании наследственной информации.
Цифрами обозначены нуклеотиды
4.2 Свойства ДНК как вещества наследственности и изменчивости
4.2.1 Самовоспроизведение наследственного материала. Репликация ДНК
Одним из основных свойств материала наследственности является его способность к самокопированию - репликация. Это свойство обеспечивается особенностями химической организации молекулы ДНК, состоящей из двух комплементарных цепей. В процессе репликации на каждой полинуклеотидной цепи материнской молекулы ДНК синтезируется комплементарная ей цепь. В итоге из одной двойной спирали ДНК образуются две идентичные двойные спирали. Такой способ удвоения молекул, при котором каждая дочерняя молекула содержит одну материнскую и одну вновь синтезированную цепь, называют полуконсервативным.
Для осуществления репликации цепи материнской ДНК должны быть отделены друг от друга, чтобы стать матрицами, на которых будут синтезироваться комплементарные цепи дочерних молекул.
Инициация репликации осуществляется в особых участках ДНК, обозначаемых ori (от англ. origin - начало). Они включают последовательность, состоящую из 300 нуклеотидных пар, узнаваемую специфическими белками. Двойная спираль ДНК в этих локусах разделяется на две цепи, при этом, как правило, по обе стороны от точки начала репликации образуются области расхождения полинуклеотидных цепей - репликационные вилки, которые движутся в противоположных от локуса ori направлениях. Между репликационными вилками образуется структура, называемая репликационным глазком, где на двух цепях материнской ДНК образуются новые полинуклеотидные цепи (рис 8, А).
С помощью фермента геликазы, разрывающего водородные связи, двойная спираль ДНК расплетается в точках начала репликации. Образующиеся при этом одинарные цепи ДНК связываются специальными дестабилизирующими белками, которые растягивают остовы цепей, делая их азотистые основания доступными для связывания с комплементарными нуклеотидами, находящимися в нуклеоплазме. На каждой из цепей, образующихся в области репликационной вилки, при участии фермента ДНК-полимеразы осуществляется синтез комплементарных цепей (рис 8, Б).

Рис.8. Область начала репликации. Репликационная вилка
А. Образование репликационного глазка.
В. Область репликационной вилки в молекуле ДНК
В процессе синтеза репликационные вилки движутся вдоль материнской спирали в противоположных направлениях, захватывая все новые зоны.
Разделение спирально закрученных цепей родительской ДНК ферментом геликазой вызывает появление супервитков перед репликационной вилкой. Это объясняется тем, что при расхождении каждых 10 пар нуклеотидов, образующих один виток спирали, родительская ДНК должна совершить один полный оборот вокруг своей оси. Следовательно, для продвижения репликационной вилки вся молекула ДНК перед ней должна была бы быстро вращаться, что потребовало бы большой затраты энергии. В действительности это не наблюдается благодаря особому классу белков, называемых ДНК-топоизомеразами. Топоизомераза разрывает одну из цепей ДНК, что дает ей возможность вращаться вокруг второй цепи. Это ослабляет накопившееся напряжение в двойной спирали ДНК (рис.9).
К высвобождающимся водородным связям нуклеотидных последовательностей разделенных родительских цепей присоединяются свободные нуклеотиды из нуклеоплазмы, где они присутствуют в виде дезоксирибонуклеозидгрифосфатов: дАТФ, дГТФ, дЦТФ, дТТФ. Комплементарный нуклеозидтрифосфат образует водородные связи с определенным основанием материнской цепи ДНК. Затем при участии фермента ДНК-полимеразы он связывается фосфодиэфирной связью с предшествующим нуклеотидом вновь синтезируемой цепи, отдавая при этом неорганический пирофосфат (рис.10).
Поскольку ДНК-полимераза присоединяет очередной нуклеотид к ОН-группе в 3'-положении предшествующего нуклеотида, цепь постепенно удлиняется на ее 3'-конце.
Особенностью ДНК-полимеразы является ее неспособность начать синтез новой полинуклеотидной цепи путем простого связывания двух нуклеозидтрифосфатов: необходим 3'-ОН-конец какой-либо полинуклеотидной цепи, спаренной с матричной цепью ДНК, к которой ДНК-полимераза может лишь добавлять новые нуклеотиды. Такую полинук-леотидную цепь называют затравкой или праймером.
Роль затравки для синтеза полинуклеотидных цепей ДНК в ходе репликации выполняют короткие последовательности РНК, образуемые при участии фермента РНК-праймазы (рис.11). Указанная особенность ДНК-полимеразы означает, что матрицей при репликации может служить лишь цепь ДНК, несущая спаренную с ней затравку, которая имеет свободный 3'-ОН-конец.

Рис.9. Разрыв одной из цепей ДНК с помощью фермента ДНК-топоизомеразы: I - ДНК-топоизомераза образует ковалентаую связь с одной из фосфатных групп ДНК (верхняя цепь); II - в результате разрыва фосфодиэфирной связи в одной полинуклеотидной цепи вокруг соответствующей ей связи другой цепи осуществляется вращение, которое снимает напряжение, вызванное расхождением двух цепей ДНК в области репликационной вилки; III - после снятия напряжения в спирали ДНК происходит спонтанное отделение ДНК-топоизомеразы и восстановление фосфодиэфирной связи в цепи ДНК
Способность ДНК-полимеразы осуществлять сборку полинуклеотида в направлении от 5' - к 3' - концу при антипараллельном соединении двух цепей ДНК означает, что процесс репликации должен протекать на них по-разному. Действительно, если на одной из матриц (3' → 5') сборка новой цепи происходит непрерывно от 5' - к 3'-концу и она постепенно удлиняется на 3'-конце, то другая цепь, синтезируемая на матрице (5' → 3'), должна была бы расти от 3' - к 5'-концу. Это противоречит направлению действия фермента ДНК-полимеразы.

Рис.10. Присоединение очередного нуклеотида к дочерней цепи ДНК, синтезируемой при участии ДНК-полимеразы: ФФ-пирофосфат
В настоящее время установлено, что синтез второй цепи ДНК осуществляется короткими фрагментами (фрагменты Оказаки) также в направлении от 5' - к 3'-концу (по типу шитья "назад иголкой"). У прокариот фрагменты Оказаки содержат от 1000 до 2000 нуклеотидов, у эукариот они значительно короче (от 100 до 200 нуклеотидов). Синтезу каждого такого фрагмента предшествует образование РНК-затравки длиной около 10 нуклеотидов. Вновь образованный фрагмент с помощью фермента ДНК-лигазы соединяется с предшествующим фрагментом после удаления его РНК-затравки (рис.12, А).
В связи с указанными особенностями репликационная вилка является асимметричной. Из двух синтезируемых дочерних цепей одна строится непрерывно, ее синтез идет быстрее и эту цепь называют лидирующей. Синтез другой цепи идет медленнее, так как она собирается из отдельных фрагментов, требующих образования, а затем удаления РНК-затравки. Поэтому такую цепь называют запаздывающей (отстающей). Хотя отдельные фрагменты образуются в направлении 5' → 3', в целом эта цепь растет в направлении 3' → 5' (рис.3.12, А). В виду того, что от локуса ori как правило начинаются две репликационные вилки, идущие в противоположных направлениях, синтез лидирующих цепей в них идет на разных цепях материнской ДНК (рис 12, Б). Конечным результатом процесса репликации является образование двух молекул ДНК, нуклеотидная последовательность которых идентична таковой в материнской двойной спирали ДНК.

Рис.11. Схема реакции синтеза короткой РНК-затравки, катализируемой РНК-праймазой
Рассмотренная последовательность событий, происходящих в ходе репликативного синтеза, предполагает участие целой системы ферментов: геликазы, топоизомеразы, дестабилизирующих белков, ДНК-полимеразы и других, совместно действующих в области репликационной вилки (рис 13).
Репликация ДНК у про- и эукариот в основных чертах протекает сходно, однако, скорость синтеза у эукариот (около 100 нуклеотидов/с) на порядок ниже, чем у прокариот (1000 нуклеотидов/с). Причиной этого может быть образование ДНК эукариот достаточно прочных соединений с белками, что затрудняет ее деспирализацию, необходимую для осуществления репликативного синтеза.
Фрагмент ДНК от точки начала репликации до точки ее окончания образует единицу репликации - репликон. Однажды начавшись в точке начала (локус on), репликация продолжается до тех пор, пока весь репликон не будет дуплицирован. Кольцевые молекулы ДНК прокариотических клеток имеют один локус on и представляют собой целиком отдельные репликоны. Эукариотические хромосомы содержат большое число репликонов. В связи с этим удвоение молекулы ДНК, расположенной вдоль эукариотической хромосомы, начинается в нескольких точках. В разных репликонах удвоение может идти в разное время или одновременно.
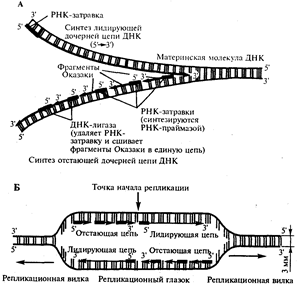
Рис. 12. Синтез двух дочерних цепей ДНК на разных цепях материнской молекулы
А. В связи с антипараллельностью цепей ДНК синтез дочерних цепей идет по-разному, на верхней материнской цепи дочерняя цель синтезируется непрерывно-лидирующая цепь, на нижней материнской цепи дочерняя цепь собирается из фрагментов Оказаки - отстающая цепь.
Б. Синтез лидирующих цепей в разнонаправленных вилках происходит на разных цепях материнской ДНК
4.2.2 Механизмы сохранения нуклеогидной последовательности ДНК. Химическая стабильность. Репликация. Репарация
Для поддержания главных характеристик клетки или организма на протяжении их жизни, а также в ряду поколений наследственный материал должен отличаться устойчивостью к внешним воздействиям или должны существовать механизмы коррекции возникающих в нем изменений. В живой природе используются оба фактора. Третьим фактором является точность копирования нуклеотидных последовательностей материнской ДНК в процессе ее репликации.

Рис.13. Белки, участвующие в процессе репликации ДНК
ДНК-геликаза расплетает двойную спираль ДНК, разделяя ее полинуклеотидные цепи; дестабилизирующие белки выпрямляют участок цепи ДНК; ДНК-топоизомераза разрывает фосфодиэфирную связь в одной из полинуглеотидных цепей ДНК, снимая напряжение, вызываемое расплетенисм спирали и расхождением цепей в репликационной вилке; РНК-праймаза синтезирует РНК-затравки для дочерней цепи и для каждого фрагмента Оказаки; ДНК-полимераза осуществляет непрерывный синтез лидирующей цепи и синтез фрагментов Оказаки отстающей цепи; ДНК-лигаза сшивает фрагменты Оказаки после удаления РНК-затравки
По реакционной способности молекулы ДНК относятся к категории химически инертных веществ. Известно, что роль вещества наследственности может выполнять не только ДНК, но и РНК (некоторые вирусы). Считают, что выбор в пользу ДНК обусловлен ее более низкой по сравнению с РНК реакционной способностью.
Рассмотренный выше механизм репликации отличается чрезвычайно высокой точностью воспроизведения структуры ДНК. При удвоении ДНК ошибки возникают в среднем с частотой 1·10-6 комплементарных пар оснований.
В поддержании высокой точности репликации важная роль принадлежит прежде всего ферменту ДНК-полимеразе. Этот фермент осуществляет отбор необходимых нуклеотидов из числа имеющихся в ядерном соке нуклеозидтрифосфатов (АТФ, ТТФ, ГТФ, ЦТФ), точное присоединение их к матричной цепи ДНК и включение в растущую дочернюю цепь. Частота включения неправильных нуклеотидов на этой стадии составляет 1·10-5 пар оснований.
Такие ошибки в работе ДНК-полимеразы связаны с возникновением измененных форм азотистых оснований, которые образуют "незаконные" пары с основаниями материнской цепи. Например, измененная форма цитозина вместо гуанина связывается водородными связями с аденином. В результате в растущую цепь ДНК включается ошибочный нуклеотид. Быстрый переход измененной формы такого основания в обычную нарушает его связывание с матрицей, появляется неспаренный 3'-ОН-конец растущей цепи ДНК. В этой ситуации включается механизм самокоррекции, осуществляемый ДНК-полимеразой (или тесно связанным с ней ферментом - редактирующей эндонуклеазой). Самокоррекция заключается в отщеплении ошибочно включенного в цепь ДНК нуклеотида, не спаренного с матрицей (рис.14). Следствием самокоррекции является снижение частоты ошибок в 10 раз (с 10-5 до 10-6 ).
Несмотря на эффективность самокоррекции, в ходе репликации после удвоения ДНК в ней обнаруживаются ошибки. Особенно часто это наблюдается при нарушении концентрации четырех нуклеозидтрифосфатов в окружающем субстрате. Значительная часть изменений возникает также в молекулах ДНК в результате спонтанно происходящих процессов, связанных с потерей пуриновых оснований - аденина и гуанина (апуринизацией) - или дезаминированием цитозина, который превращается в урацил. Частота последних изменений достигает 100 на 1 геном/сут.
Содержащиеся в ДНК основания могут изменяться под влиянием реакционноспособных соединений, нарушающих их нормальное спаривание, а также под действием ультрафиолетового излучения, которое может вызвать образование ковалентной связи между двумя соседними остатками тимина в ДНК (димеры тимина). Названные изменения в очередном цикле репликации должны привести либо к выпадению пар оснований в дочерней ДНК, либо к замене одних пар другими. Указанные изменения действительно сопровождают каждый цикл репликации ДНК, однако их частота значительно меньше, чем должна была бы быть. Это объясняется тем, что большинство изменений такого рода устраняется благодаря действию механизма репарации (молекулярного восстановления) исходной нуклеотидной последовательности ДНК.
Механизм репарации основан на наличии в молекуле ДНК двух комплементарных цепей. Искажение последовательности нуклеотидов в одной из них обнаруживается специфическими ферментами. Затем соответствующий участок удаляется и замещается новым, синтезированным на второй комплементарной цепи ДНК. Такую репарацию называют эксцизионной, т.е. с "вырезанием" (рис.15). Она осуществляется до очередного цикла репликации, поэтому ее называют также дорепликативной.

Рис.14. Схема процесса коррекции при синтезе ДНК:
I-включение в цепь ДНК нуклеотида с измененной (таутомерной) формой цитоэина, который "незаконно" спаривается с аденином; II - быстрый переход цитозина в обычную форму нарушает его спаривание с аденином; неспаренный 3'-ОН-конец синтезируемой цепи препятствует дальнейшему ее удлинению под действием ДНК-полимеразы; III - ДНК-полимераза удаляет незаконный нуклеотид, в результате чего вновь появляется спаренный с матрицей 3 '-ОН-конец; IV - ДНК-полимераза продолжает наращивание цепи на 3'-ОН-конце.
Восстановление исходной структуры ДНК требует участия ряда ферментов. Важным моментом в запуске механизма репарации является обнаружение ошибки в структуре ДНК. Нередко такие ошибки возникают во вновь синтезированной цепи в процессе репликации. Ферменты репарации должны обнаружить именно эту цепь. У многих видов живых организмов вновь синтезированная цепь ДНК отличается от материнской степенью метилирования ее азотистых оснований, которое отстает от синтеза. Репарации при этом подвергается неметилированная цепь. Объектом узнавания ферментами репарации могут также служить разрывы в цепи ДНК. У высших организмов, где синтез ДНК происходит не непрерывно, а отдельными репликонами, вновь синтезируемая цепь ДНК имеет разрывы, что делает возможным ее узнавание. Восстановление структуры ДНК при утрате пуриновых оснований одной из ее цепей предполагает обнаружение дефекта с помощью фермента эндонуклеазы, которая разрывает фосфоэфирную связь в месте повреждения цепи. Затем измененный участок с несколькими примыкающими к нему нуклеотидами удаляется ферментом экзонуклеазой, а на его месте в соответствии с порядком оснований комплементарной цепи образуется правильная нуклеотидная последовательность (рис.15).

Рис.15. Схема эксцизионной, дорепликативной репарации ДНК.
При изменении одного из оснований в цепи ДНК в восстановлении исходной структуры принимают участие ферменты ДНК-гликозилазы числом около 20. Они специфически узнают повреждения, обусловленные дезаминированием, алкилированием и другими структурными преобразованиями оснований. Такие модифицированные основания удаляются. Возникают участки, лишенные оснований, которые репарируются, как при утрате пуринов. Если восстановление нормальной структуры не осуществляется, например в случае дезаминирования азотистых оснований, происходит замена одних пар комплементарных оснований другими - пара Ц-Г может заменяться парой Т-А и т.п. .
Образование в полинуклеотидных цепях под действием УФ-лучей тиминовых димеров (Т-Т) требует участия ферментов, узнающих не отдельные измененные основания, а более протяженные повреждения структуры ДНК. Репаративный процесс в этом случае также связан с удалением участка, несущего димер, и восстановлением нормальной последовательности нуклеотидов путем синтеза на комплементарной цепи ДНК.
В том случае, когда система эксцизионной репарации не исправляет изменения, возникшего в одной цепи ДНК, в ходе репликации происходит фиксация этого изменения и оно становится достоянием обеих цепей ДНК. Это приводит к замене одной пары комплементарных нуклеотидов на другую либо к появлению разрывов (брешей) во вновь синтезированной цепи против измененных участков. Восстановление нормальной структуры ДНК при этом может произойти и после репликации.
Пострепликативная репарация осуществляется путем рекомбинации (обмена фрагментами) между двумя вновь образованными двойными спиралями ДНК. Примером такой пострепликативной репарации может служить восстановление нормальной структуры ДНК при возникновении тиминовых димеров (Т-Т), когда они не устраняются самопроизвольно под действием видимого света (световая репарация) или в ходе дорепликативной эксцизионной репарации.
Ковалентные связи, возникающие между рядом стоящими остатками тимина, делают их не способными к связыванию с комплементарными нуклеотидами. В результате во вновь синтезируемой цепи ДНК появляются разрывы (бреши), узнаваемые ферментами репарации. Восстановление целостности новой полинуклеотидной цепи одной из дочерних ДНК осуществляется благодаря рекомбинации с соответствующей ей нормальной материнской цепью другой дочерней ДНК. Образовавшийся в материнской цепи пробел заполняется затем путем синтеза на комплементарной ей полинуклеотидной цепи (рис.16). Проявлением такой пострепликативной репарации, осуществляемой путем рекомбинации между цепями двух дочерних молекул ДНК, можно считать нередко наблюдаемый обмен материалом между сестринскими хроматидами (рис.17).
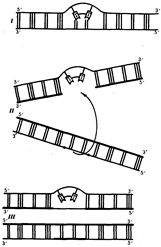
Рис.16. Схема пострепликативной репарации ДНК:
I - возникновение тиминового димера в одной из цепей ДНК;
II - образование "бреши" во вновь синтезируемой цепи против измененного участка материнской молекулы после репликации (стрелкой показано последующее заполнение "бреши" участком из соответствующей цепи второй дочерней молекулы ДНК);
III - восстановление целостности дочерней цепи верхней молекулы за счет рекомбинации и в нижней молекуле за счет синтеза на комплементарной цепи

Рис.17. Межхроматидные обмены (указаны стрелками)
В ходе дорепликативной и пострепликативной репарации восстанавливается большая часть повреждений структуры ДНК. Однако, если в наследственном материале клетки возникает слишком много повреждений и часть из них не ликвидируется, включается система индуцируемых (побуждаемых) ферментов репарации (SOS-система). Эти ферменты заполняют бреши, восстанавливая целостность синтезируемых полинуклеотидных цепей без точного соблюдения принципа комплементарности. Вот почему иногда сами процессы репарации могут служить источником стойких изменений в структуре ДНК (мутаций). Названная реакция также относится к SOS-системе.
Если в клетке, несмотря на осуществляемую репарацию, количество повреждений структуры ДНК остается высоким, в ней блокируются процессы репликации ДНК. Такая клетка не делится, а значит, не передает возникших изменений потомству.
Вызываемая повреждениями ДНК остановка клеточного цикла в сочетании с невозможностью молекулярной репарации измененного наследственного материала может с участием белка, синтез которого контролируется геном р53, приводить к активации процесса самоликвидации (апотпоз) дефектной клетки с целью устранения ее из организма.
Таким образом, обширный набор различных ферментов репарации осуществляет непрерывный "осмотр" ДНК, удаляя из нее поврежденные участки и способствуя поддержанию стабильности наследственного материала. Совместное действие ферментов репликации (ДНК-полимераза и редактирующая эндонуклеаза) и ферментов репарации обеспечивает достаточно низкую частоту ошибок в молекулах ДНК, которая поддерживается на уровне 1 · 10-9 пар измененных нуклеотидов на геном. При размере генома человека 3 · 109 нуклеотидных пар это означает появление около 3 ошибок на реплицирующийся геном. Вместе с тем даже этот уровень достаточен для образования за время существования жизни на Земле значительного генетического разнообразия в виде генных мутаций.
4.2.3 Изменения нуклеотидных последовательностей ДНК .
Нескорректированные изменения химической структуры генов, воспроизводимые в последовательных циклах репликации и проявляющиеся у потомства в виде новых вариантов признаков, называют генными мутациями.
Изменения структуры ДНК, образующей ген, можно разделить на три группы. Мутации первой группы заключаются в замене одних оснований другими. Они составляют около 20% спонтанно возникающих генных изменений. Вторая группа мутаций обусловлена сдвигом рамки считывания, происходящим при изменении количества нуклеотидных пар в составе гена. Наконец, третью группу представляют мутации, связанные с изменением порядка нуклеотидных последовательностей в пределах гена (инверсии).
Мутации по типу замены азотистых оснований. Эти мутации происходят в силу ряда конкретных причин. Одной из них может быть возникающее случайно или под влиянием конкретных химических агентов изменение структуры основания, уже включенного в спираль ДНК. Если такая измененная форма основания остается не замеченной ферментами репарации, то при ближайшем цикле репликации она может присоединять к себе другой нуклеотид. Примером может служить дезаминирование цитозина, превращающегося в урацил самопроизвольно или под влиянием азотистой кислоты (рис.18). Образующийся при этом урацил, не замеченный ферментом ДНК-гликозилазой, при репликации соединяется с аденином, который впоследствии присоединяет тимидиловый нуклеотид. В результате пара Ц-Г замещается в ДНК парой Т-А (рис. 19, I). Дезаминирование метилированного цитозина превращает его в тимин (см. рис.3.18). Тимидиловый нуклеотид, являясь естественным компонентом ДНК, не обнаруживается ферментами репарации как изменение и при следующей репликации присоединяет адениловый нуклеотид. В результате вместо пары Ц-Г в молекуле ДНК также появляется пара Т-А (рис. 19, II).

Рис.18. Спонтанное дезаминирование цитозина
Другой причиной замены оснований может быть ошибочное включение в синтезируемую цепь ДНК нуклеотида, несущего химически измененную форму основания или его аналог. Если эта ошибка остается не замеченной ферментами репликации и репарации, измененное основание включается в процесс репликации, что нередко приводит к замене одной пары на другую. Примером этого может служить присоединение в ходе репликации к аденину материнской цепи нуклеотида с 5-бромурацилом (5-БУ), аналогичного тимидиловому нуклеотиду. При последующей репликации 5-БУ охотнее присоединяет к себе не аденин, а гуанин. Гуанин в ходе дальнейшего удвоения образует комплементарную пару с цитозином. В итоге пара А-Т заменяется в молекуле ДНК парой Г-Ц (рис. 20).

Рис. 19. Мутации по типу замены основания (дезаминирование азотистых оснований в цепи ДНК):
I - превращение цитозина в урацил, замена Ц-Г-пары на Т-А-пару;
II - превращение метил-цитозина в тимин, замена Ц-Г-пары на Т-А-пару
Из приведенных примеров видно, что изменения структуры молекулы ДНК по типу замены оснований возникают либо до, либо в процессе репликации первоначально в одной полинуклеотидной цепи. Если такие изменения не исправляются в ходе репарации, то при последующей репликации они становятся достоянием обеих цепей ДНК.

Рис. 20. Мутации по типу замены оснований (включение аналога азотистого основания при репликации ДНК)
Следствием замены одной пары комплементарных нуклеотидов на другую является образование нового триплета в нуклеотидной последовательности ДНК, кодирующей последовательность аминокислот в пептидной цепи. Это может и не отразиться на структуре пептида в том случае, если новый триплет будет "синонимом" прежнего, т.е. будет кодировать ту же аминокислоту. Например, аминокислота валин шифруется четырьмя триплетами: ЦАА, ЦАГ, ЦАТ, ЦАЦ. Замена третьего основания в любом из этих триплетов не изменит его смысла (вырожденность генетического кода).
В том случае, когда вновь возникший триплет шифрует другую аминокислоту, изменяются структура пептидной цепи и свойства соответствующего белка. В зависимости от характера и места случившейся замены специфические свойства белка изменяются в разной степени. Известны случаи, когда замена лишь одной аминокислоты в пептиде существенно влияет на свойства белка, что проявляется в изменении более сложных признаков. Примером может служить изменение свойств гемоглобина человека при серповидно-клеточной анемии (рис.21). В таком гемоглобине- (HbS) (в отличие от нормального НbА) - в р-глобиновых цепях в шестом положении глутаминовая кислота заменена валином. Это является следствием замены одного из оснований в триплете, шифрующем глутаминовую кислоту (ЦТТ или ЦТЦ). В результате появляется триплет, шифрующий валин (ЦАТ или ЦАЦ). В данном случае замена одной аминокислоты в пептиде существенно изменяет свойства глобина, входящего в состав гемоглобина (снижается его способность связываться с 02), у человека развиваются признаки серповидно-клеточной анемии.
В некоторых случаях замена одного основания на другое может привести к появлению одного из нонсенс-триплетов (АТТ, АТЦ, АЦТ), не шифрующего никакой аминокислоты. Последствием такой замены будет прерывание синтеза пептидной цепи. Подсчитано, что замены нуклеотидов в одном триплете приводят в 25% случаев к образованию триплетов-синонимов; в 2-3 - бессмысленных триплетов, в 70 - 75% - к возникновению истинных генных мутаций.
Таким образом, мутации по типу замены оснований могут возникать как в результате спонтанных изменений структуры основания в одной из цепей уже существующей двойной спирали ДНК, так и в ходе репликации во вновь синтезируемой цепи. В том случае, если эти изменения не исправляются в процессе репарации (или, наоборот, возникают в ходе репарации), они фиксируются в обеих цепях и далее будут воспроизводиться в следующих циклах репликации. Следовательно, важным источником возникновения таких мутаций являются нарушения процессов репликации и репарации.
Мутации со сдвигом рамки считывания. Этот тип мутаций составляет значительную долю спонтанных мутаций. Они происходят вследствие выпадения или вставки в нуклеотидную последовательность ДНК одной или нескольких пар комплементарных нуклеотидов. Большая часть изученных мутаций, вызывающих сдвиг рамки, обнаружена в последовательностях, состоящих из одинаковых нуклеотидов.
Изменению числа нуклеотидных пар в цепи ДНК способствуют воздействия на генетический материал некоторых химических веществ, например акридиновых соединений. Деформируя структуру двойной спирали ДНК, они приводят к вставке дополнительных оснований или их выпадению при репликации. Примером служат мутации, полученные у фага Т4 при воздействии профлавина. Они состоят во включении или удалении всего одной нуклеотидной пары. Важной причиной изменения количества нуклеотидных пар в гене по типу крупных делений (выпадений) может быть рентгеновское облучение. У плодовой мухи, например, известна мутация гена, контролирующего окраску глаза, которая вызывается облучением и состоит в делении порядка 100 нуклеотидных пар.

Рис.21. Плейотропный эффект замены одной аминокислоты в β-цепи гемоглобина человека, приводящей к развитию серповидно-клеточной анемии
Большое число мутаций по типу вставок происходит вследствие включения в последовательность нуклеотидов подвижных генетических элементов - транспозонов. Транспозоны - это достаточно протяженные нуклеотидные последовательности, встроенные в геномы эу - и прокариотических клеток, способные самопроизвольно менять свое положение. С определенной вероятностью вставки и делении могут возникать в результате ошибок рекомбинации при неравноценном внутригенном кроссинговере (рис.22).

Рис.22. Мутации со сдвигом рамки считывания (неравноценный обмен при внутригенном кроссинговере):
I - разрывы аллельпых генов в разных участках и обмен фрагментами между ними;
II - выпадение 3-й и 4-й пар нуклеотидов, сдвиг рамки считывания;
III - удвоение 3-й и 4-й пар нуклеотидов, сдвиг рамки считывания
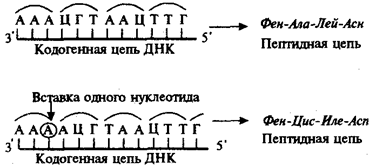
Рис.23. Следствие изменения количества нуклеотидных пар в молекуле ДНК
Сдвиг рамки считывания в результате вставки одного нуклеотида в кодогенную цепь приводит к изменению состава зашифрованного в ней пептида
При непрерывности считывания и неперекрываемости генетического кода изменение количества нуклеотидов, как правило, приводит к сдвигу рамки считывания и изменению смысла биологической информации, записанной в данной последовательности ДНК (рис.23). Однако, если количество вставленных или утраченных нуклеотидов кратно трем, сдвига рамки может не произойти, но это приведет к включению дополнительных аминокислот или выпадению части их из полипептидной цепи. Возможным следствием сдвига рамки является возникновение нонсенстриплетов, ведущее к синтезу укороченных пептидных цепей.
Мутации по типу инверсии нуклеотидных последовательностей в гене. Данный тип мутаций происходит вследствие поворота участка ДНК на 180°. Обычно этому предшествует образование молекулой ДНК петли, в пределах которой репликация идет в направлении, обратном правильному.
В пределах инвертированного участка нарушается считывание информации, в результате изменяется аминокислотная последовательность белка.
4.2.4 Элементарные единицы изменчивости генетического материала. Мутон. Рекон
Ген представляет собой элементарную единицу функции наследственного материала. Это означает, что фрагмент молекулы ДНК, соответствующий отдельному гену и определяющий благодаря содержащейся в нем биологической информации возможность развития конкретного признака, является далее неделимым в функциональном отношении. Сведения о генных мутациях, изложенные выше, указывают на значение изменений химической структуры, затрагивающих не весь ген, а отдельные его участки, вследствие чего появляются новые варианты признака.
Минимальное количество наследственного материала, способное, изменяясь, приводить к появлению вариантов признака, соответствует элементарной единице мутационного процесса и называется мутоном. Рассмотренные выше примеры генных мутаций свидетельствуют о том, что достаточно заменить одну пару комплементарных оснований в гене, чтобы изменились свойства кодируемого им белка. Таким образом, мутон соответствует одной паре комплементарных нуклеотидов.
Часть генных мутаций по типу вставок и выпадений нуклеотидных пар происходит вследствие неравноценного обмена между молекулами ДНК при кроссинговере, т.е. при нарушении рекомбинации между ними. Это сопровождается сдвигом рамки считывания и приводит к нарушению синтеза пептидной цепи с заданными свойствами. Наблюдения показывают, что для искажения записанной в гене биологической информации достаточно вставки или выпадения одной пары нуклеотидов. Из сказанного следует, что элементарная единица рекомбинации - рекон - на молекулярном уровне соответствует одной паре нуклеотидов.
Возникающие самопроизвольно или под влиянием различных внешних воздействий изменения нуклеотидных последовательностей приводят к тому, что один и тот же ген может существовать в нескольких вариантах, различающихся по содержащейся в них биологической информации. Конкретную форму существования гена, определяющую возможность развития конкретного варианта данного признака, называют аллелем. Аллели гена располагаются в одном и том же участке-локусе-определенной хромосомы, которая в норме может одновременно содержать лишь один из серии аллелей. Это делает аллели альтернативными (взаимоисключающими) вариантами существования гена.
Изменения химической структуры могут возникать в различных участках гена. Если они совместимы с жизнью, т.е. не приводят к гибели клеток или организмов - носителей данных мутаций, все они сохраняются в генофонде вида.
Присутствие в генофонде вида одновременно различных аллелей гена называют множественным аллелизмом. Примером этому служат разные варианты окраски глаз у плодовой мухи: белая, вишневая, красная, абрикосовая, эозиновая, - обусловленные различными аллелями соответствующего гена. У человека, как и у других представителей органического мира, множественный аллелизм свойствен многим генам. Так, три аллеля гена I определяют групповую принадлежность крови по системе АВ0 (IA , IB , I0 ). Два аллеля имеет ген, обусловливающий резус-принадлежность. Более ста аллелей насчитывают гены α - и β-полипептидов гемоглобина.
Причиной множественного аллелизма являются случайные изменения структуры гена (мутации), сохраняемые в процессе естественного отбора в генофонде популяции. Многообразие аллелей, рекомбинирующихся при половом размножении, определяет степень генотипического разнообразия среди представителей данного вида, что имеет большое эволюционное значение, повышая жизнеспособность популяций в меняющихся условиях их существования. Кроме эволюционного и экологического значения аллельное состояние генов оказывает большое влияние на функционирование генетического материала. В диплоидных соматических клетках эукариотических организмов большинство генов представлено двумя аллелями, которые совместно влияют на формирование признаков.
4.2.5 Функциональная классификация генных мутаций
Изменения структуры гена, как правило, являются неблагоприятными, снижая жизнеспособность клетки, организма (вредные мутации), и иногда приводят к их гибели (летальные мутации). Реже возникающие мутации существенно не отражаются на жизнеспособности их носителей, поэтому их рассматривают как нейтральные. Наконец, крайне редко возникают аллели, оказывающие благоприятное действие (полезные мутации), обеспечивая их носителям преимущественное выживание. В большинстве случаев вновь возникший аллель гена выступает как рецессивный по отношению к распространенному в природе аллелю "дикого" типа, т.е. не проявляется в сочетании с ним. Но иногда мутантная форма гена может быть доминантной, т.е. подавлять проявление "дикого" аллеля, который чаще встречается в генофонде популяции.
4.2.6 Механизмы, снижающие неблагоприятный эффект генных мутаций
В результате генных мутаций изменяется смысл биологической информации. Последствия этого могут быть двоякого рода. В условиях обитания, изменяющихся незначительно, новая информация обычно снижает выживаемость. При резкой смене условий существования, при освоении новой экологической ниши наличие разнообразной информации полезно. В связи с этим интенсивность мутационного процесса в природных условиях поддерживается на уровне, не вызывающем катастрофического снижения жизнеспособности вида. Важная роль в ограничении неблагоприятных последствий мутаций принадлежит антимутационным механизмам, возникшим в эволюции.
Некоторые из этих механизмов рассмотрены выше. Речь идет об особенностях функционирования ДНК-полимеразы, отбирающей требуемые нуклеотиды в процессе репликации ДНК, а также осуществляющей самокоррекцию при образовании новой цепи ДНК наряду с редактирующей эндонуклеазой. Подробно разобраны различные механизмы репарации структуры ДНК, роль вырожденности генетического кода. Решением этой задачи служит триплетность биологического кода, которая допускает минимальное число замен внутри триплета, ведущих к искажению информации. Так, 64% замен третьего нуклеотида в триплетах не дает изменения их смыслового значения. Правда, замены второго нуклеотида в 100% приводят к искажению смысла триплета.
Фактором защиты против неблагоприятных последствий генных мутаций служит парность хромосом в диплоидном кариотипе соматических клеток эукариот.
Парность аллелей генов препятствует фенотипическому проявлению мутаций, если они имеют рецессивный характер.
Определенный вклад в снижение вредных последствий генных мутаций вносит явление экстракопирования генов, кодирующих жизненно важные макромолекулы. Оно заключается в наличии в генотипе нескольких десятков, а иногда и сотен идентичных копий таких генов. Примером могут служить гены рРНК, тРНК, гистоновых белков, без которых жизнедеятельность любой клетки невозможна.
При наличии экстракопий мутационное изменение в одном или даже нескольких одинаковых генах не ведет к катастрофическим для клетки последствиям. Копий, остающихся неизменными, вполне достаточно, чтобы обеспечить нормальное функционирование.
Существенное значение имеет также функциональная неравнозначность замен аминокислот в полипептиде. Если новая и сменяемая аминокислоты сходны по физико-химическим свойствам, изменения третичной структуры и биологических свойств белка незначительны.
Так, мутантные гемоглобины HbS и НЬС человека отличаются от нормального гемоглобина НЬА заменой в 6-м положении р-цепи глутаминовой кислоты соответственно на валин или лизин. Первая замена резко изменяет свойства гемоглобина и приводит к развитию тяжелого заболевания - серповидно-клеточной анемии.
При второй замене свойства гемоглобина изменяются в гораздо меньшей степени.
Причиной этих различий является то, что глутаминовая кислота и лизин проявляют сходные гидрофильные свойства, тогда как валин - это гидрофобная аминокислота.
Таким образом, перечисленные механизмы способствуют сохранению отобранных в ходе эволюции генов и одновременно накоплению в генофонде популяции различных их аллелей, формируя резерв наследственной изменчивости. Последний определяет высокую эволюционную пластичность популяции, т.е. способность выживать в разнообразных условиях.
4.3 Использование генетической информации в процессах жизнедеятельности
4.3.1 Роль РНК в реализации наследственной информации
Наследственная информация, записанная с помощью генетического кода, хранится в молекулах ДНК и размножается для того, чтобы обеспечить вновь образуемые клетки необходимыми "инструкциями" для их нормального развития и функционирования. Вместе с тем непосредственного участия в жизнеобеспечении клеток ДНК не принимает. Роль посредника, функцией которого является перевод наследственной информации, сохраняемой в ДНК, в рабочую форму, играют рибонуклеиновые кислоты - РНК.
В отличие от молекул ДНК рибонуклеиновые кислоты представлены одной полинуклеотидной цепью, которая состоит из четырех разновидностей нуклеотидов, содержащих сахар, рибозу, фосфат и одно из четырех азотистых оснований - аденин, гуанин, урацил или цитозин. РНК синтезируется на молекулах ДНК при помощи ферментов РНК-полимераз с соблюдением принципа комплементарности и антипараллельности, причем аденину ДНК в РНК комплементарен урацил. Все многообразие РНК, действующих в клетке, можно разделить на три основных вида: мРНК, тРНК, рРНК.
Матричная, или информационная, РНК (мРНК, или иРНК). Транскрипция. Для того чтобы синтезировать белки с заданными свойствами, к месту их построения поступает "инструкция" о порядке включения аминокислот в пептидную цепь. Эта инструкция заключена в нуклеотидной последовательности матричных, или информационных РНК (мРНК, иРНК), синтезируемых на соответствующих участках ДНК. Процесс синтеза мРНК называют транскрипцией.
Синтез мРНК начинается с обнаружения РНК-полимеразой особого участка в молекуле ДНК, который указывает место начала транскрипции - промотора. После присоединения к промотору РНК-полимераза раскручивает прилежащий виток спирали ДНК. Две цепи ДНК в этом месте расходятся, и на одной из них фермент осуществляет синтез мРНК. Сборка рибонуклеотидов в цепь происходит с соблюдением их комплементарности нуклеотидам ДНК, а также антипараллельно по отношению к матричной цепи ДНК. В связи с тем, что РНК-полимераза способна собирать полинуклеотид лишь от 5'-конца к 3'-концу, матрицей для транскрипции может служить только одна из двух цепей ДНК, а именно та, которая обращена к ферменту своим 3'-концом (3' → 5'). Такую цепь называют кодогенной (рис.3.24). Антипараллельность соединения двух полинуклеотидных цепей в молекуле ДНК позволяет РНК-полимеразе правильно выбрать матрицу для синтеза мРНК.
Продвигаясь вдоль кодогенной цепи ДНК, РНК-полимераза осуществляет постепенное точное переписывание информации до тех пор, пока она не встречает специфическую нуклеотидную последовательность - терминатор транскрипции. В этом участке РНК-полимераза отделяется как от матрицы ДНК, так и от вновь синтезированной мРНК (рис.25). Фрагмент молекулы ДНК, включающий промотор, транскрибируемую последовательность и терминатор, образует единицу транскрипции - транскриптон.
В процессе синтеза, по мере продвижения РНК-полимеразы вдоль молекулы ДНК, пройденные ею одноцепочечные участки ДНК вновь объединяются в двойную спираль. Образуемая в ходе транскрипции мРНК содержит точную копию информации, записанной в соответствующем участке ДНК. Тройки рядом стоящих нуклеотидов мРНК, шифрующие аминокислоты, называют кодонами. Последовательность кодонов мРНК шифрует последовательность аминокислот в пептидной цепи. Кодонам мРНК соответствуют определенные аминокислоты (табл.1).
Таблица 1. Генетический код мРНК (подчеркнуты кодоны-терминаторы). Второй нуклеотид
| У | Ц | А | Г | ||||||||
П Е Р В Ы Й Н У К Л Е О Т И Д |
У |
УУУ УУЦ УУФ УУГ |
Фен Лей |
УЦУ УЦЦ УЦА УЦГ |
Сер |
УАУ УАЦ УАА УАГ |
Тир |
УГУ УГЦ УГА УГГ |
Цис Три |
У Ц А Г |
Т Р Е Т И Й Н У К Л Е О Т И Д |
Ц |
ЦУУ ЦУЦ ЦУА ЦУГ |
Лей |
ЦЦУ ЦЦЦ ЦЦА ЦЦГ |
Про |
ЦАУ ЦАЦ ЦАА ЦАГ |
Гис Глн |
ЦГУ ЦГЦ ЦГА ЦГГ |
Арг |
У Ц А Г |
||
А |
АУУ АУЦ АУА АУГ |
Иле Мет |
АЦУ АЦЦ АЦА АЦГ |
Тре |
ААУ ААЦ ААА ААГ |
Асн Лиз |
АГУ АГЦ АГА АГГ |
Сер Арг |
У Ц А Г |
||
Г |
ГУУ ГУЦ ГУА ГУГ |
Вал |
ГЦУ ГЦЦ ГЦА ГЦГ |
Ала |
ГАУ ГАЦ ГАА ГАГ |
Лея Глу |
ГГУ ГГЦ ГГА ГГГ |
Гли |
У Ц А Г |
||

Рис.24. Схема синтеза мРНК
Матрицей для транскрипции мРНК служит кодогенная цепь ДНК, обращенная к ферменту своим 3-концом
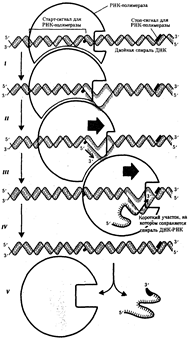
Рис. 25. Роль РНК-полимеразы в транскрипции:
I - обнаружение промоторной области в молекуле ДНК и раскручивание спирали ДНК; II - инициация синтеза цепи РНК путем связывания двух первых рибонуклеозидгрифосфатов; III - наращивание цепи РНК в направлении 5' → 3' путем присоединения рибонуклеозидгрифосфатов; IV - высвобождение 5'-конца синтезируемой РНК и восстановление двойной спирали ДНК; V - окончание синтеза РНК в области терминатора, отделение полимеразы от завершенной цепи РНК
Транспортная РНК (тРНК). Трансляция. Важная роль в процессе использования наследственной информации клеткой принадлежит транспортной РНК (тРНК). Доставляя необходимые аминокислоты к месту сборки пептидных цепей, тРНК выполняет функцию трансляционного посредника.
Молекулы тРНК представляют собой полинуклеотидные цепи, синтезируемые на определенных последовательностях ДНК. Они состоят из относительно небольшого числа нуклеотидов - 75-95. В результате комплементарного соединения оснований, которые находятся в разных участках полинуклеотидной цепи тРНК, она приобретает структуру, напоминающую по форме лист клевера (рис.26).

Рис.26. Строение типичной молекулы тРНК
В ней выделяют четыре главные части, выполняющие различные функции. Акцепторный "стебель" образуется двумя комплементарно соединенными концевыми частями тРНК. Он состоит из семи пар оснований.3'-конец этого стебля несколько длиннее и формирует одноцепочечный участок, который заканчивается последовательностью ЦЦА со свободной ОН-группой. К этому концу присоединяется транспортируемая аминокислота. Остальные три ветви представляют собой комплементарно спаренные последовательности нуклеотидов, которые заканчиваются неспаренными участками, образующими петли. Средняя из этих ветвей - антикодоновая - состоит из пяти пар нуклеотидов и содержит в центре своей петли антикодон. Антикодон - это три нуклеотида, комплементарные кодону мРНК, который шифрует аминокислоту, транспортируемую данной тРНК к месту синтеза пептида.
Между акцепторной и антикодоновой ветвями располагаются две боковые ветви. В своих петлях они содержат модифицированные основания - дигидроуридин (D-петля) и триплет TψC, где у - псевдоуриаин (Т^С-петля). Между аитикодоновой и Т^С-ветвями содержится дополнительная петля, включающая от 3-5 до 13-21 нуклеотидов.
В целом различные виды тРНК характеризуются определенным постоянством нуклеотидной последовательности, которая чаще всего состоит из 76 нуклеотидов. Варьирование их числа связано главным образом с изменением количества нуклеотидов в дополнительной петле. Комплементарные участки, поддерживающие структуру тРНК, как правило, консервативны. Первичная структура тРНК, определяемая последовательностью нуклеотидов, формирует вторичную структуру тРНК, имеющую форму листа клевера. В свою очередь, вторичная структура обусловливает трехмерную третичную структуру, для которой характерно образование двух перпендикулярно расположенных двойных спиралей (рис.27). Одна из них образована акцепторной и ТψС-ветвями, другая - антикодоновой и D-ветвями.
На конце одной из двойных спиралей располагается транспортируемая аминокислота, на конце другой - антикодон. Эти участки оказываются максимально удаленными друг от друга. Стабильность третичной структуры тРНК поддерживается благодаря возникновению дополнительных водородных связей между основаниями полинуклеотидной цепи, находящимися в разных ее участках, но пространственно сближенных в третичной структуре.
Различные виды тРНК имеют сходную третичную структуру, хотя и с некоторыми вариациями.

Рис.27. Пространственная организация тРНК:
I - вторичная структура тРНК в виде "клеверного листа", определяемая ее первичной структурой (последовательностью нуклеотидов в цепи);
II - двумерная проекция третичной структуры тРНК;
III - схема укладки молекулы тРНК в пространстве
Одной из особенностей тРНК является наличие в ней необычных оснований, возникающих вследствие химической модификации уже после включения нормального основания в полинуклеотидную цепь. Эти измененные основания обусловливают большое структурное многообразие тРНК при общем плане их строения. Наибольший интерес представляют модификации оснований, формирующих антикодон, которые влияют на специфичность его взаимодействия с кодоном. Например, нетипичное основание инозин, иногда стоящий в 1-м положении антикодона тРНК, способен комплементарно соединяться с тремя разными третьими основаниями кодона мРНК - У, Ц и А (рис.3.28). Так как одной из особенностей генетического кода является его вырожденность (см. разд.3.4.1.2), многие аминокислоты шифруются несколькими кодонами, которые, как правило, различаются своим третьим основанием. Благодаря неспецифичности связывания модифицированного основания антикодона одна тРНК узнает несколько кодонов-синонимов.

Рис.28. Соединение инозина водородными связями с тремя различными азотистыми основаниями Водородные связи обозначены точками
Установлено также существование нескольких видов тРНК, способных соединяться с одним и тем же кодоном. В результате в цитоплазме клеток встречается не 61 (по количеству кодонов), а около 40 различных молекул тРНК. Этого количества достаточно, чтобы транспортировать 20 разных аминокислот к месту сборки белка.
Наряду с функцией точного узнавания определенного кодона в мРНК молекула тРНК осуществляет доставку к месту синтеза пептидной цепи строго определенной аминокислоты, зашифрованной с помощью данного кодона. Специфическое соединение тРНК со "своей" аминокислотой протекает в два этапа и приводит к образованию соединения, называемого аминоацил-тРНК (рис.29).
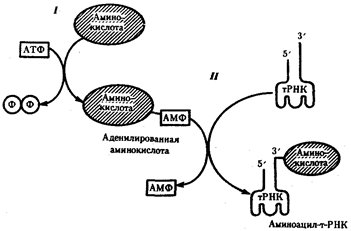
Рис.29. Присоединение аминокислоты к соответствующей тРНК:
I - 1-й этап, взаимодействие аминокислоты и АТФ с выделением пирофосфата;
II - 2-й этап, присоединение адепилированной аминокислоты к 3'-концу РНК
На первом этапе аминокислота активируется, взаимодействуя своей карбоксильной группой с АТФ. В результате образуется адепилированная аминокислота.
На втором этапе это соединение взаимодействует с ОН-группой, находящейся на 3'-конце соответствующей тРНК, и аминокислота присоединяется к нему своей карбоксильной группой, высвобождая при этом АМФ. Таким образом, этот процесс протекает с затратой энергии, получаемой при гидролизе АТФ до АМФ.
Специфичность соединения аминокислоты и тРНК, несущей соответствующий антикодон, достигается благодаря свойствам фермента аминоацил-тРНК-синтетазы. В цитоплазме существует целый набор таких ферментов, которые способны к пространственному узнаванию, с одной стороны, своей аминокислоты, а с другой - соответствующего ей антикодона тРНК (рис.3.30). Наследственная информация, "записанная" в молекулах ДНК и "переписанная" на мРНК, расшифровывается в ходе трансляции благодаря двум процессам специфического узнавания молекулярных поверхностей. Сначала фермент аминоацил-тРНК-синтетаза обеспечивает соединение тРНК с транспортируемой ею аминокислотой. Затем аминоацил-тРНК комплементарно спаривается с мРНК благодаря взаимодействию антикодона с кодоном. С помощью системы тРНК язык нуклеотидной цепи мРНК. транслируется в язык аминокислотной последовательности пептида (рис.30).
Рибосомная РНК (рРНК). Рибосомный цикл синтеза белка. Процесс взаимодействия мРНК и тРНК, обеспечивающий трансляцию информации с языка нуклеотидов на язык аминокислот, осуществляется на рибосомах. Последние представляют собой сложные комплексы рРНК и разнообразных белков, в которых первые образуют каркас. Рибосомные РНК являются не только структурным компонентом рибосом, но и обеспечивают связывание их с определенной нуклеотидной последовательностью мРНК. Этим устанавливаются начало и рамка считывания при образовании пептидной цепи. Кроме того, они обеспечивают взаимодействие рибосомы и тРНК. Многочисленные белки, входящие в состав рибосом наряду с рРНК, выполняют как структурную, так и ферментативную роль.

Рис.30. Схема трансляции генетического кода: I - присоединение аминокислоты (триптофана) к соответствующей тРНК с помощью фермента аминоацил-тРНК-синтетазы; II - присоединение тРНК, несущей свою аминокислоту, к мРНК благодаря связыванию ее антикодона с кодоном мРНК
Рибосомы про- и эукариот очень сходны по структуре и функциям. Они состоят из двух субчастиц: большой и малой. У эукариот малая субчастица образована одной молекулой рРНК и 33 молекулами разных белков. Большая субчастица объединяет три молекулы рРНК и около 40 белков. Прокариотические рибосомы и рибосомы митохондрий и пластид содержат меньше компонентов.
В рибосомах имеется две бороздки. Одна из них удерживает растущую полипептидную цепь, другая - мРНК. Кроме того, в рибосомах выделяют два участка, связывающих тРНК. В аминоацильном, А-участке размещается аминоацил-тРНК, несущая определенную аминокислоту. В пептидильном, П-участке располагается обычно тРНК, которая нагружена цепочкой аминокислот, соединенных пептидными связями. Образование А - и П-участков обеспечивается обеими субчастицами рибосомы.
В каждый момент рибосома экранирует сегмент мРНК протяженностью около 30 нуклеотидов. При этом обеспечивается взаимодействие только двух тРНК с двумя расположенными рядом кодонами мРНК (рис.31).
Трансляция информации на "язык" аминокислот выражается в постепенном наращивании пептидной цепи в соответствии с инструкцией, заключенной в мРНК. Этот процесс протекает на рибосомах, которые обеспечивают последовательность расшифровки информации с помощью тРНК. В ходе трансляции можно выделить три фазы: инициацию, элонгацию и терминацию синтеза пептидной цепи.

Рис.31. Участки связывания молекул тРНК и рибосомы:
I - ненагруженная рибосома, II - нагруженная рибосома; ак - аминокислота
Фаза инициации, или начало синтеза пептида, заключается в объединении двух находящихся до этого порознь в цитоплазме субчастиц рибосомы на определенном участке мРНК и присоединении к ней первой аминоацил-тРНК. Этим задается также рамка считывания информации, заключенной в мРНК (рис.32).
В молекуле любой мРНК вблизи ее 5'-конца имеется участок, комплементарный рРНК малой субчастицы рибосомы и специфически узнаваемый ею. Рядом с ним располагается инициирующий стартовый кодон АУТ, шифрующий аминокислоту метионин. Малая субчастица рибосомы соединяется с мРНК таким образом, что стартовый кодон АУТ располагается в области, соответствующей П-участку. При этом только инициирующая тРНК, несущая метионин, способна занять место в недостроенном П-участке малой субчастицы и комплементарно соединиться со стартовым кодоном. После описанного события происходит объединение большой и малой субчастиц рибосомы с образованием ее пептидильного и аминоацильного участков (рис.3.32).

Рис.32. Инициация белкового синтеза:
I - соединение малой субчаспщы рибосомы с мРНК, присоединение к стартовому кодону несущей метионин тРНК, которая располагается в недостроенном П-участке; II - соединение большой и малой субчастиц рибосомы с образованием П - и А-участков; следующий этап связан с размещением в А-участке аминоацил-тРНК, соответствующей расположенному в нем кодону мРНК,-начало элонгации; ак - аминокислота
К концу фазы инициации П-участок занят аминоацил-тРНК, связанной с метионином, тогда как в А-участке рибосомы располагается следующий за стартовым кодон.
Описанные процессы инициации трансляции катализируются особыми белками - факторами инициации, которые подвижно связаны с малой субчастицей рибосомы. По завершении фазы инициации и образования комплекса рибосома - мРНК - инициирующая аминоацил-тРНК эти факторы отделяются от рибосомы.
Фаза элонгации, или удлинения пептида, включает в себя все реакции от момента образования первой пептидной связи до присоединения последней аминокислоты. Она представляет собой циклически повторяющиеся события, при которых происходит специфическое узнавание аминоацил-тРНК очередного кодона, находящегося в А-участке, комплементарное взаимодействие между антикодоном и кодоном.
Благодаря особенностям трехмерной организации тРНК при соединении ее антикодона с кодоном мРНК. транспортируемая ею аминокислота располагается в А-участке, поблизости от ранее включенной аминокислоты, находящейся в П-участке. Между двумя аминокислотами образуется пептидная связь, катализуемая особыми белками, входящими в состав рибосомы. В результате предыдущая аминокислота теряет связь со своей тРНК и присоединяется к аминоацил-тРНК, расположенной в А-участке. Находящаяся в этот момент в П-участке тРНК высвобождается и уходит в цитоплазму (рис.33). Перемещение тРНК, нагруженной пептидной цепочкой, из А-участка в П-участок сопровождается продвижением рибосомы по мРНК на шаг, соответствующий одному кодону. Теперь следующий кодон приходит в контакт с А-участком, где он будет специфически "опознан" соответствующей аминоацил-тРНК, которая разместит здесь свою аминокислоту. Такая последовательность событий повторяется до тех пор, пока в А-участок рибосомы не поступит кодон-терминатор, для которого не существует соответствующей тРНК.

Рис.33. Фаза элонгации в синтезе белка:
1-й этап - аминоацил-тРНК присоединяется к кодону, расположенному в А-участке;
2-й этап - между аминокислотами, расположенными в А - и П-участках, образуется пептидиая связь: тРНК, расположенная в П-участке, освобождается от своей аминокислоты и покидает рибосому;
3-й этап - рибосома перемещается по мРНК на один кодон так, что тРНК, нагруженная пептидной цепочкой, переходит из А-участка в П-участок; свободный А-участок может быть занят соответствующей аминоацил-тРНК
Рис.34. Терминация синтеза пептидной цепи:
1-й этап - присоединение фактора освобождения к стоп-кодону;
2-й этап - терминация, высвобождение завершенного пептида;
3-й этап - диссоциация рибосомы на две субчастицы
Сборка пептидной цепи осуществляется с достаточно большой скоростью, зависящей от температуры. У бактерий при 37 °С она выражается в добавлении к подипептиду от 12 до 17 аминокислот в 1 с. В эукариотических клетках эта скорость ниже и выражается в добавлении двух аминокислот в 1 с.
Фаза терминации, или завершения синтеза полипептида, связана с узнаванием специфическим рибосомным белком одного из терминирующих кодонов (УАА, УАГ или У ГА), когда тот входит в зону А-участка рибосомы. При этом к последней аминокислоте в пептидной цепи присоединяется вода, и ее карбоксильный конец отделяется от тРНК. В результате завершенная пептидная цепь теряет связь с рибосомой, которая распадается на две субчастицы (рис.34).
4.3.2 Особенности организации и экспрессии генетической информации у про- и эукариот
По химической организации материала наследственности и изменчивости эукариотические и прокариотические клетки принципиально не отличаются друг от друга. Генетический материал у них представлен ДНК. Общим для них является и принцип записи генетической информации, а также генетический код. Одни и те же аминокислоты шифруются у про - и эукариот одинаковыми кодонами. Принципиально одинаковым образом у названных типов клеток осуществляется и использование наследственной информации, хранящейся в ДНК. Сначала она транскрибируется в нуклеотидную последовательность молекулы мРНК, а затем транслируется в аминокислотную последовательность пептида на рибосомах с участием тРНК. Однако некоторые особенности организации наследственного материала, отличающие эукариотические клетки от прокариотических, обусловливают различия в использовании их генетической информации.
Наследственный материал прокариотической клетки содержится главным образом в единственной кольцевой молекуле ДНК. Она располагается непосредственно в цитоплазме клетки, где также находятся необходимые для экспрессии генов тРНК и ферменты, часть из которых заключена в рибосомах. Гены прокариот состоят целиком из кодирующих нуклеотидных последовательностей, реализующихся в ходе синтеза белков, тРНК или рРНК.
Наследственный материал эукариот больше по объему, чем у прокариот. Он расположен в основном в особых ядерных структурах - хромосомах, которые отделены от цитоплазмы ядерной оболочкой. Необходимый для синтеза белков аппарат, состоящий из рибосом, тРНК, набора аминокислот и ферментов, находится в цитоплазме клетки.
Значительные отличия имеются в молекулярной организации генов эукариотической клетки. В большинстве из них кодирующие последовательности экзоны прерываются интронными участками, которые не используются при синтезе тРНК, рРНК или пептидов. Количество таких участков варьирует в разных генах. Установлено, что ген овальбумина кур включает 7 интронов, а ген проколлагена млекопитающих - 50. Эти участки удаляются из первично-транскрибируемой РНК, в связи с чем использование генетической информации в эукариотической клетке происходит несколько иначе. В прокариотической клетке, где наследственный материал и аппарат биосинтеза белка пространственно не разобщены, транскрипция и трансляция происходят почти одновременно. В эукариотической клетке эти два этапа не только пространственно отделены ядерной оболочкой, но и во времени их разделяют процессы созревания мРНК, из которой должны быть удалены неинформативные последовательности (рис.35).

Рис. 35. Обобщенная схема процесса экспрессии генетической информации в эукариотической клетке
Кроме указанных различий на каждом этапе экспрессии генетической информации можно отметить некоторые особенности течения этих процессов у про - и эукариот.
Транскрипция у про - и эукариот. Транскрипция - это синтез РНК на матрице ДНК. У прокариот синтез всех трех видов РНК катализируется одним сложным белковым комплексом - РНК-полимеразой.
Транскрипционный аппарат эукариотических клеток включает три ядерные РНК-полимеразы, а также РНК-полимеразы митохондрий и пластид. РНК-полимераза I обнаруживается в ядрышках клеток и отвечает за транскрипцию генов рРНК. РНК-полимераза II локализуется в ядерном соке и отвечает за синтез предшественника мРНК. РНК-полимераза III - небольшая фракция, находящаяся в ядерном соке и осуществляющая синтез малых рРНК и тРНК. Каждый из этих ферментов имеет две большие субъединицы и до 10 малых. РНК-полимеразы митохондрий и пластид отличаются от ядерных.
Ферментный комплекс РНК-полимеразы специфически узнает некую нуклеотидную последовательность (часто не одну), расположенную на определенном расстоянии от стартовой точки транскрипции, - промотор. Стартовой точкой считают нуклеотид ДНК, которому соответствует первый нуклеотид, включаемый ферментом в РНК-транскрипт.
У прокариот обычно недалеко от стартовой точки против хода транскрипции располагается последовательность из шести нуклеотидов - ТАТААТ, называемая блоком Прибнова. Это среднестатистическая последовательность, состоящая из наиболее часто встречаемых оснований, самыми консервативными из которых являются 1,2 и 6-е основания. Наличие в этой последовательности оснований, преимущественно соединенных двойными водородными связями с комплементарными основаниями другой цепи, очевидно, облегчает локальное плавление двойной спирали ДНК и образование двух ее одноцепочечных участков при контакте с РНК-полимеразой. Блок Прибнова располагается в положении от - 11 до - 5 или от - 14 до - 8, т.е. за несколько нуклеотидов перед стартовой точкой транскрипции (рис.36). Обнаруживая эту последовательность, РНК-полимераза прочно связывается с ней и начинает синтез РНК. Столь же важная роль в установлении контакта РНК-полимеразы с ДНК принадлежит другой нуклеотидной последовательности, центр которой находится в положении - 35. Ее называют областью узнавания - ТТГАЦА. Между двумя указанными участками расстояние достаточно постоянно и составляет от 16 до 19 пар нуклеотидов (п. н).
Промоторы эукариотических генов также включают по меньшей мере две специфические нуклеотидные последовательности, центры которых находятся в положении - 25 и - 75 п. н.
На расстоянии 19-27 нуклеотидов от стартовой точки против хода транскрипции у многих генов эукариот обнаружена среднестатистическая последовательность ТАТА Т АА Т (ТАТА-блок, или блок Хогнесса), в которой, так же как в блоке Прибнова у прокариот, преобладают основания, образующие более слабые связи. Вторую последовательность, встречаемую во многих промоторах эукариот и состоящую из ГГЦ Т ЦААТЦТ, обозначают как ЦААТ-блок. Она занимает положение между - 70 и - 80 нуклеотидами и также является областью, узнаваемой полимеразой. В некоторых генах обнаружены многокомпонентные промоторы.
Так, в отдельных генах вируса герпеса для эффективной инициации транскрипции необходимы три последовательности ДНК, расположенные между - 19 и - 27, между - 47 и - 61, а также между - 80 и - 105 нуклеотидами.
Рис.36. Точки контакта для РНК-полимеразы, находящиеся в верхней цепи ДНК (промотор)
Особенности промоторных участков свидетельствуют о том, что для инициации транскрипции имеет значение не только сочетание оснований в определенных областях промотора, но и взаимное расположение в молекуле ДНК этих областей, с которыми связывается ферментный комплекс РНК-полимеразы.
После установления контакта между РНК-полимеразой и промоторным участком начинается сборка молекулы РНК, в которую первым чаще всего включается нуклеотид, несущий пуриновое основание (как правило, аденин) и содержащий три 5'-фосфатных остатка.
Далее, по мере продвижения РНК-полимеразы вдоль молекулы ДНК происходит постепенное удлинение цепи РНК, которое продолжается до встречи фермента с областью терминатора. Терминатор - это участок, где прекращается дальнейший рост цепи РНК и происходит ее освобождение от матрицы ДНК. РНК-полимераза также отделяется от ДНК, которая восстанавливает свою двухцепочечную структуру.

Рис.37. Область ДНК с двойной симметрией - палиндром:
I - палиндром, в котором имеется последовательность, одинаковая при чтении в противоположных направлениях;
II - палиндром, в котором заштрихованный инвертированный повтор находится на расстоянии от оси симметрии
В прокариотических клетках терминаторы обязательно содержат палиндромы - двухцепочечные последовательности нуклеотидов ДНК, которые одинаково читаются в обоих направлениях (рис.37). Участок РНК, транскрибированный с такой последовательности, способен образовывать двухцепочечные шпильки за счет комплементарного спаривания нуклеотидов палиндрома. Возможно, это и является сигналом для завершения транскрипции, узнаваемым РНК-полимеразой (рис.3.38). Возникающие шпильки, видимо, останавливают полимеразу на терминаторе. Следом за шпилькой в молекулу РНК включается последовательность из нуклеотидов, содержащих урацил (полиУ), которая, вероятно, принимает участие в высвобождении РНК от матрицы ДНК. Действительно, полиУ-последовательность РНК, соединенная с полиадениловой (полиА) последовательностью ДНК, характеризуется слабым взаимодействием. Обращает на себя внимание тот факт, что участок ДНК, богатый парами А-Т, встречается не только в месте инициации транскрипции (блок Прибнова), но и в терминаторной области.
Бактериальные терминаторы значительно различаются по своей эффективности. Некоторые из них как бы не замечаются РНК-полимеразой, и она продолжает транскрипцию за пределами терминатора. Такое прочитывание терминатора при транскрипции бактериальных генов наблюдается в результате предотвращения терминации специфическими белками - факторами антитерминации. Следствием антитерминации является синтез полицистронной мРНК, включающей в себя информацию, списанную с нескольких последовательно расположенных структурных генов.
Терминаторы эукарйогических генов изучены в меньшей степени, чем у проскариот, но в них также обнаружены районы, богатые Г-Ц парами, соединенными тройными водородными связями, в которых располагается, участок с А-Т парами. На этом участке в транскрипт включается полиУ-последовательность, слабо взаимодействующая с матричной полиА-областыо ДНК.
Возможно, область терминатора, богатая Г-Ц парами, играет определенную роль в остановке РНК-полимеразы, а участок РНК, содержащий УУУУ обеспечивает отделение транскрипта от матрицы ДНК.
У эукариот не обнаружено образования структур, подобных шпилькам в прокариотических РНК. Поэтому, каким образом у них осуществляется терминация транскрипции, остается неясным.
В составе всех мРНК можно выделить кодирующие участки, представляющие набор кодонов, которые шифруют последовательность аминокислот в пептиде. Как правило, эти участки начинаются стартовым кодоном АУГ, но иногда у бактерий используется кодон ГУТ. На конце кодирующей последовательности располагается терминирующий кодон. Помимо кодирующих участков в мРНК на обоих концах могут располагаться дополнительные последовательности. На 5'-конце это лидерный участок, расположенный перед стартовым кодоном. На 3'-конце - трейлер, следующий за кодоном-терминатором.
Рис.38. Образование шпильки участком РНК при терминации транскрипции у прокариот
Область РНК, несущая палиндром, образует комплементарно спаривающуюся структуру - шпильку (инвертированные повторы заштрихованы)
В полицистронной мРНК прокариот между кодирующими участками имеются межцистронные области, варьирующие по размерам (рис.3.39).

Рис.39. Полицистронная матричная РНК прокариот:
1 - некодирующие области, 2 - межцистронные области, 3 - кодирующие области, 4 - терминирующие кодоны
В связи с тем что прокариотические гены целиком состоят из нуклеотидных последовательностей, участвующих в кодировании информации, транскрибированные с них РНК сразу после их синтеза способны выполнять функцию матриц для трансляции. Лишь в исключительных случаях требуется их предварительное созревание - процессинг.
В отличие от прокариотических генов большинство генов эукариотических клеток прерывисты, так как несут в своем составе неинформативные нуклеотидные последовательности - интроны, не участвующие в кодировании информации. В связи с этим первичные транскрипты, синтезированные РНК-полимеразой II, обладают большими, чем необходимо для трансляции, размерами и оказываются менее стабильными. В совокупности они образуют так называемую гетерогенную ядерную РНК (тРНК), которая прежде чем выйти из ядра и начать активно функционировать в цитоплазме, подвергается процессингу и превращается в зрелые мРНК.
Процессинг эукариотических мРНК. Созревание, или процессинг, мРНК предполагает модифицирование первичного транскрипта и удаление из него некодирующих интронных участков с последующим соединением (сплайсингом) кодирующих последовательностей - экзонов. Модифицирование первичного транскрипта эукариотической мРНК начинается вскоре после синтеза его 5'-конца, содержащего одно из пуриновых оснований (аденин или гуанин). На этом конце образуется колпачок - кэп, который блокирует 5'-конец мРНК путем присоединения к первому нуклеотиду транскрипта трифосфонуклеозида, содержащего гуанин, связью 5'-5'.
Гффф + фффАфN… → ГфффАфN. + фф + ф В результате образуется последовательность ГфффАфЧМ..., в которой остаток туанина находится в обратной ориентации по отношению к другим нуклеотидам мРНК. Модификация 5'-конца мРНК предполагает также метилирование присоединенного гуанина и первых двух-трех оснований первичного транскрипта (рис.3.40). Образуемые на 5' - концах мРНК кэпы обеспечивают узнавание молекул мРНК малыми субчастицами рибосом в цитоплазме. Кэширование осуществляется еще до окончания синтеза первичного транскрипта.

Рис. 40. Образование зрелой мРНК эукариот в ходе процессинга:
1 - некодирующие последовательности, 2 - экзоны, 3 - интроны, 4 - кодон-терминатор
После завершения транскрипции происходит удаление части нуклеотидов на 3'-конце первичного транскрипта и присоединение к нему последовательности, состоящей из 100-200 остатков адениловой кислоты (полиА) (рис.3.40). Считают, что эта последовательность способствует дальнейшему процессингу и транспорту зрелой мРНК из ядра. После выхода мРНК в цитоплазму ее полиА-последовательность постепенно укорачивается под действием ферментов, отщепляющих нуклеотиды на 3'-конце. Таким образом, по длине полиА-последовательности можно косвенно судить о времени пребывания мРНК в цитоплазме. Возможно, добавление полиА-последовательности в ходе процессинга повышает стабильность мРНК. Однако около трети мРНК вообще не содержат полиА-участка. К ним относятся, например, гистоновые мРНК.
Образование кэпа на 5'-конце и полиА-последовательности на 3'-конце характерно только для процессинга РНК, синтезируемых РНК-полимеразой II. Кроме метилирования при формировании кэпов в мРНК высших эукариот происходит метилирование небольшой части внутренних нуклеотидов с частотой приблизительно одно на тысячу оснований мРНК.
Наряду с модифицированием мРНК эукариот процессинг предполагает удаление из первичных транскриптов неинформативных для данного белка интронных участков, размер которых варьирует от 100 до 10 000 нуклеотидов и более. На долю интронов приходится около 80% всей гяРНК. Удаление интронов с последующим соединением экзонных участков называют сплайсингом (рис.40).
Сплайсинг представляет собой механизм, который должен обеспечивать удаление из первичного транскрипта строго определенных интронных участков. Нарушение этого процесса может привести к сдвигу рамки считывания при трансляции и невозможности синтеза нормального пептида. Закономерность вырезания интронов, очевидно, обеспечивается благодаря наличию на их концах специфических нуклеотидных последовательностей, служащих сигналами для сплайсинга.
В настоящее время описано несколько вероятных механизмов сплайсинга, обеспечивающих точность этого процесса. Возможно, она достигается действием каких-то ферментов, специфически узнающих концевые участки интронов и катализирующих разрыв фосфодиэфирных связей на границе экзон - интрон, а затем образование связей между двумя экзонами.
Установлено активное участие в сплайсинге особых малых, ядерных РНК (мяРНК), образующих комплексы с белками (мяРНП). Очевидно, мяРНК своими нуклеотидными последовательностями комплементарно взаимодействуют с концевыми участками интронов, которые образуют при этом замкнутые петли. Расщепление РНК в устье интронной петли приводит к удалению неинформативной последовательности и соединению (сплайсингу) сближенных концов экзонов.
Обсуждается также автокаталитическая способность РНК-транскрипта к сплайсингу. Описанные способы сплайсинга свидетельствуют об отсутствии универсального механизма этого процесса, однако во всех случаях достигается точное удаление интронов с образованием определенной мРНК, обеспечивающей синтез необходимого клетке белка.
В настоящее время доказана возможность альтернативного (взаимоисключающего) сплайсинга, при котором из одного и того же первичного транскрипта могут удаляться разные нуклеотидные последовательности и образовываться разные зрелые мРНК. В результате одна и та же последовательность нуклеотидов ДНК может служить информацией для синтеза разных пептидов. Альтернативный сплайсинг, вероятно, очень характерен в системе генов иммуноглобулинов у млекопитающих, где он позволяет формировать на основе одного транскрипта мРНК для синтеза разных видов антител.
Благодаря преобразованиям, происходящим с РНК-транскриптом в ходе процессинга, зрелые мРНК эукариот характеризуются большей стабильностью по сравнению с прокариотическими мРНК.
По завершении процессинга зрелая мРНК проходит отбор перед выходом в цитоплазму, куда попадает всего 5% гяРНК. Остальная часть расщепляется, не покидая ядра.
Таким образом, преобразования первичных транскриптов эукариотических генов, обусловленные их экзонитронной организацией и необходимостью перехода мРНК из ядра в цитоплазму, определяют особенности реализации генетической информации в эукариотической клетке.
Трансляция у про - и эукариот. В прокариотических клетках процесс трансляции сопряжен с синтезом мРНК: они происходят практически одновременно. В значительной степени это связано с недолговечностью бактериальной мРНК, которая достаточно быстро подвергается распаду. Взаимосвязанность транскрипции и трансляции у бактерии проявляется в согласованности скоростей этих процессов. При 37°С транскрипция идет со скоростью 2500 нуклеотидов/мин (14 кодонов/с), а трансляция осуществляется со скоростью 15 аминокислот/с.
Трансляция у прокариот начинается вскоре после образования 5'-конца мРНК, раньше, чем заканчивается ее синтез. В результате вслед за РНК-полимеразой по мРНК движутся рибосомы, осуществляющие сборку пептидных цепей (рис.41). Через некоторое время после начала транскрипции (около 1 мин) и до завершения трансляции 3'-конца матрицы начинается деградация ее 5'-конца. Ввиду того что время жизни разных мРНК не одинаково, количество белка, синтезированного на разных матрицах, различно.
Одной из особенностей трансляции у прокариот является включение в пептидную цепь в качестве первой аминокислоты модифицированного метионина - формилметионина, с которого начинаются все вновь синтезированные пептиды. Даже в том случае, когда роль стартового кодона выполняет кодом ГУГ, в обычных условиях шифрующий валин, в первом положении пептида оказывается формилметионин. Стартовый кодон АУГ или ГУГ следует за лидерным участком, который экранируется рибосомой в момент инициации трансляции.
Соединение рибосомы с мРНК обусловлено комплементарным взаимодействием нуклеотидов одной из рРНК с нуклеотидной последовательностью лидера мРНК.
Эта последовательность (Шайна-Дальгарно) располагается на расстоянии 4-7 оснований перед кодоном АУГ и обнаруживается повсеместно в лидерных участках у прокариот.
При соединении 5'-конца мРНК с малой субчастицей рибосомы стартовый кодон обычно оказывается почти в середине экранированного рибосомой фрагмента мРНК, в области, соответствующей ее П-участку.
У эукариот трансляция осуществляется в цитоплазме, куда попадает из ядра зрелая мРНК. Копированный конец мРНК распознается малой субчастицей рибосомы, затем лидирующая последовательность, содержащая до 100 нуклеотидов, взаимодействует с рРНК. При этом стартовый кодон АУГ оказывается в недостроенном П-участке рибосомы. После присоединения к стартовому кодону аминоацил-тРНК, несущей метионин, происходит воссоединение двух субчастиц рибосомы и формируются ее А - и П-участки. Синтез белка в эукариотической клетке, осуществляемый на моноцистронной мРНК, завершается после прохождения рибосомой по всей мРНК, вплоть до узнавания ею кодона-терминатора, прекращающего образование пептидных связей.
Посттрансляционные преобразования белков. Синтезированные в ходе трансляции пептидные цепи на основе своей первичной структуры приобретают вторичную и третичную, а многие - и четвертичную организацию, образуемую несколькими пептидными цепями. В зависимости от функций, выполняемых белками, их аминокислотные последовательности могут претерпевать различные преобразования, формируя функционально активные молекулы белка.
Многие мембранные белки синтезируются в виде пребелков, имеющих на N-конце лидерную последовательность, которая обеспечивает himузнавание мембраны. Эта последовательность отщепляется при созревании и встраивании белка в мембрану. Секреторные белки также имеют на N-конце лидерную последовательность, которая обеспечивает их транспорт через мембрану. Некоторые белки сразу после трансляции несут дополнительные аминокислотные пропоследовательности, определяющие стабильность предшественников активных белков. При созревании белка они удаляются, обеспечивая переход неактивного пробелка в активный белок. Например, инсулин вначале синтезируется как препроинсулин. Во время секреции пре-последовательность отщепляется, а затем проинсулин подвергается модификации, при которой из него удаляется часть цепи и он превращается в зрелый инсулин.

Рис.41. Транскрипция, трансляция и деградация мРНК у прокариот:
I - РНК-полимераза связывается с ДНК и начинает синтезировать мРНК в направлении 5' → 3';
II - по мере продвижения РНК-полимеразы к 5'-концу мРНК прикрепляются рибосомы, начинающие синтез белка;
III - группа рибосом следует за РНК-полимеразой, на 5'-конце мРНК начинается ее деградация;
IV - процесс деградации протекает медленнее, чем транскрипция и трансляция;
V - после окончания транскрипции мРНК освобождается от ДНК, на ней продолжается трансляция и деградация на 5'-конце
Формируя третичную и четвертичную организацию в ходе посттрансляционных преобразований, белки приобретают способность активно функционировать, включаясь в определенные клеточные структуры и осуществляя ферментативные и другие функции.
Рассмотренные особенности реализации генетической информации в про - и эукариотических клетках обнаруживают принципиальное сходство этих процессов. Следовательно, механизм экспрессии генов, связанный с транскрипцией и последующей трансляцией информации, которая зашифрована с помощью биологического кода, сложился в целом еще до того, как были сформированы эти два типа клеточной организации. Дивергентная эволюция геномов про - и эукариот привела к возникновению различий в организации их наследственного материала, что не могло не отразиться и на механизмах его экспресии.
Постоянное совершенствование наших знаний об организации и функционировании материала наследственности и изменчивости обусловливает эволюцию представлений о гене как функциональной единице этого материала.
Похожие рефераты:
Литература - Другое (книга по генетике)
Продукты рекомбинации: характеристика и манипулирование
Аппарат экспрессии генов и его логика
Проблемы использования генетически модифицированных организмов
Молекулы генетического аппарата
Получение рекомбинантного аденовируса CELO
Структурная и молекулярная организация генного вещества
Характеристика общих свойств микроорганизмов
Достижения генной инженерии и биотехнологии
Роль материнского генома в развитии потомка
Ответы к государственным экзаменам для эколого-биологического факультета ПетрГУ