| Похожие рефераты | Скачать .docx | Скачать .pdf |
Реферат: Синтез ДНК, РНК и белков
Тема сегодняшней лекции – синтез ДНК, РНК и белков. Синтез ДНК называется репликацией или редупликацией (удвоением), синтез РНК – транскрипцией (переписывание с ДНК), синтез белка, проводимый рибосомой на матричной РНК называется трансляцией, то есть переводим с языка нуклеотидов на язык аминокислот.
Мы постараемся дать краткий обзор всех этих процессов, в то же время останавливаясь более подробно на молекулярных деталях, для того чтобы вы получили представление, на какую глубину этот предмет изучен.
Репликация ДНК
Молекула ДНК, состоящая из двух спиралей, удваивается при делении клетки. Удвоение ДНК основано на том, что при расплетении нитей к каждой нити можно достроить комплементарную копию, таким образом получая две нити молекулы ДНК, копирующие исходную.
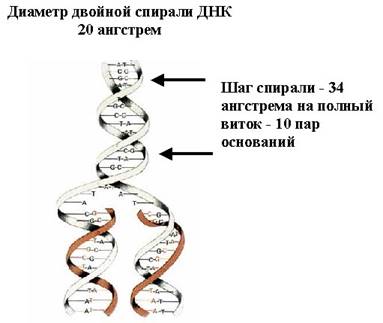
Здесь также указан один из параметров ДНК, это шаг спирали, на каждый полный виток приходится 10 пар оснований, заметим, что один шаг – это не между ближайшими выступами, а через один, так как у ДНК есть малая бороздка и большая. Через большую бороздку с ДНК взаимодействуют белки, которые распознают последовательность нуклеотидов. Шаг спирали равен 34 ангстрем, а диаметр двойной спирали – 20 ангстрем.
Репликацию ДНК осуществляет фермент ДНК-полимераза. Этот фермент способен наращивать ДНК только на 3΄– конце. Вы помните, что молекула ДНК антипараллельна, разные ее концы называются 3΄-конец и 5΄ - конец. При синтезе новых копий на каждой нити одна новая нить удлиняется в направлении от 5΄ к 3΄ , а другая – в направлении от 3΄ к 5-концу. Однако 5΄ конец ДНК-полимераза наращивать не может. Поэтому синтез одной нити ДНК, той, которая растет в "удобном" для фермента направлении, идет непрерывно (она называется лидирующая или ведущая нить), а синтез другой нити осуществляется короткими фрагментами (они называются фрагментами Оказаки в честь ученого, который их описал). Потом эти фрагменты сшиваются, и такая нить называется запаздывающей, в целом репликация этой нити идет медленней. Структура, которая образуется во время репликации, называется репликативной вилкой.
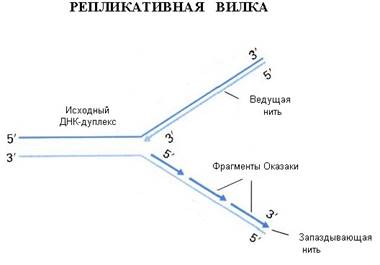
Если мы посмотрим в реплицирующуюся ДНК бактерии, а это можно наблюдать в электронном микроскопе, мы увидим, что у нее вначале образуется "глазок", затем он расширяется, в конце концов вся кольцевая молекула ДНК оказывается реплицированной. Процесс репликации происходит с большой точностью, но не абсолютной. Бактериальная ДНК-полимераза делает ошибки, то есть вставляет не тот нуклеотид, который был в матричной молекуле ДНК, примерно с частотой 10-6. У эукариот ферменты работают точнее, так как они более сложно устроены, уровень ошибок при репликации ДНК у человека оценивается как 10-7 – 10 -8 . Точность репликации может быть разной на разных участках геном, есть участки с повышенной частотой мутаций и есть участки более консервативные, где мутации происходят редко. И в этом следует различать два разных процесса: процесс появления мутации ДНК и процесс фиксации мутации. Ведь если мутации ведут к летальному исходу, они не проявятся в следующих поколениях, а если ошибка не смертельна, она закрепится в следующих поколениях, и мы сможем ее проявление наблюдать и изучить. Еще одной особенностью репликации ДНК является то, что ДНК-полимераза не может начать процесс синтеза сама, ей нужна «затравка». Обычно в качестве такой затравки используется фрагмент РНК. Если речь идет о геноме бактерии, то там есть специальная точка называемая origin (исток, начало) репликации, в этой точке находится последовательность, которая распознается ферментом, синтезирующим РНК. Он относится к классу РНК-полимераз, и в данном случае называется праймазой. РНК-полимеразы не нуждаются в затравках, и этот фермент синтезирует короткий фрагмент РНК – ту самую «затравку», с которой начинается синтез ДНК.
Транскрипция
Следующий процесс – транскрипция. На нем остановимся подробнее.
Транскрипция – синтез РНК на ДНК, то есть синтез комплементарной нити РНК на молекуле ДНК осуществляется ферментом РНК-полимеразой. У бактерий, например, кишечной палочки – одна РНК-полимераза, и все бактериальные ферменты очень похожи друг на друга ; у высших организмов (эукариотов) – несколько ферментов, они называются РНК-полимераза I, РНК-полимераза II, РНК-полимераза III, они также имеют сходство с бактериальными ферментами, но устроены сложнее, в их состав входит больше белков. Каждый вид эукариотической РНК-полимеразы обладает своими специальными функциями, то есть транскрибирует определенный набор генов. Нить ДНК, которая служит матрицей для синтеза РНК при транскрипции называется смысловой или матричной. Вторая нить ДНК называется некодирующей (комплементарная ей РНК не кодирует белки, она "бессмысленная").
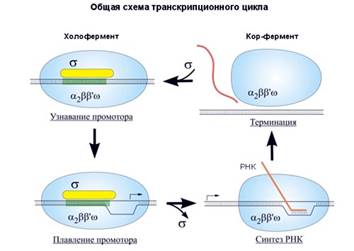
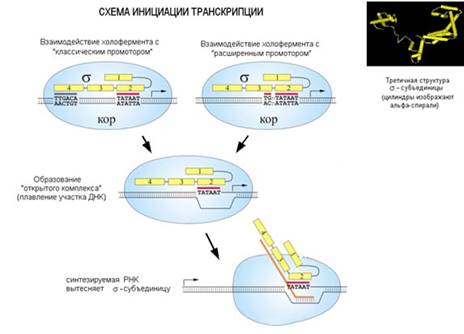
В процессе транскрипции можно выделить три этапа. Первый этап - инициация транскрипции – начало синтеза нити РНК, образуется первая связь между нуклеотидами. Затем идет наращивание нити, ее удлинение – элонгация, и, когда синтез завершен, происходит терминация, освобождение синтезированной РНК. РНК-полимераза при этом «слезает» с ДНК и готова к новому циклу транскрипции. Бактериальная РНК-полимераза изучена очень подробно. Она состоит из нескольких белковых-субъединиц: двух α-субъединиц (это маленькие субъединицы), β- и β΄-субъединиц (большие субъединицы) и ω-субъединицы. Вместе они образуют так называемый минимальный фермент, или кор-фермент. К этому кор-ферменту может присоединяться σ-субъединица. σ-субъединица необходима для начала синтеза РНК, для инициации транскрипции. После того, как инициация осуществилась, σ-субъединица отсоединяется от комплекса, и дальнейшую работу (элонгацию цепи) ведет кор-фермент. При присоединении к ДНК σ-субъединица распознает участок, на котором должна начинаться транскрипция. Он называется промотор. Промотор - это последовательность нуклеотидов, указывающих на начало синтеза РНК. Без σ-субъединицы кор-фермент промотор распознать не может. σ-субъединица вместе с кор-ферментом называется полным ферментом, или холоферментом.
Связавшись с ДНК, а именно с промотором, который распознала σ-субъединица, холофермент расплетает двунитевую спираль и начинает синтез РНК. Участок расплетенной ДНК – это точка инициации транскрипции, первый нуклеотид, к которому должен комплементарно быть присоединен рибонуклеотид. Инициируется транскрипция, σ-субъединица уходит, а кор-фермент продолжает элонгацию цепи РНК. Затем происходит терминация, кор-фермент освобождается и становится готов к новому циклу синтеза.
Как происходит элонгация транскрипции?
РНК наращивается на 3΄-конце. Присоединением каждого нуклеотида кор-фермент делает шаг по ДНК и сдвигается на один нуклеотид. Так как все в мире относительно, то можно сказать, что кор-фермент неподвижен, а сквозь него «протаскивается» ДНК. Понятно, что результат будет таким же. Но мы будем говорить о движении по молекуле ДНК. Размер белкового комплекса, составляющего кор-фермент, 150 Ǻ. Размеры РНК-полимеразы - 150×115×110Ǻ. То есть это такая наномашина. Скорость работы РНК-полимеразы – до 50 нуклеотидов в секунду. Комплекс кор-фермента с ДНК и РНК называется элонгационным комплексом. В нем находится ДНК-РНК гибрид. То есть это участок, на котором ДНК спарена с РНК, и 3΄-конец РНК открыт для дальнейшего роста. Размер этого гибрида – 9 пар оснований. Расплетенный участок ДНК занимает примерно 12 пар оснований.
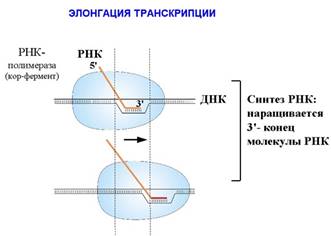
РНК-полимераза связанна с ДНК перед расплетенным участком. Этот участок называется передним дуплексом ДНК, его размер – 10 пар оснований. Полимераза связана также с более длинной частью ДНК, называемой задним дуплексом ДНК. Размер матричных РНК, которые синтезируют РНК-полимеразы у бактерий, могут достигать 1000 нуклеотидов и больше. В эукариотических клетках размер синтезируемых ДНК может достигать 100000 и даже нескольких миллионов нуклеотидов. Правда, неизвестно, существуют ли они в таких размерах в клетках, или в процессе синтеза они могут успеть процессировать.
Элонгационный комплекс довольно стабилен, т.к. он должен выполнить большую работу. То есть, сам по себе он с ДНК не «свалится». Он способен перемещаться по ДНК со скоростью до 50 нуклеотидов в секунду. Этот процесс называется перемещение (или, транслокация). Взаимодействие ДНК с РНК-полимеразой (кор-ферментом) не зависит от последовательности этой ДНК, в отличие от σ-субъединицы. И кор-фермент при прохождении определенных сигналов терминации завершает синтез ДНК.
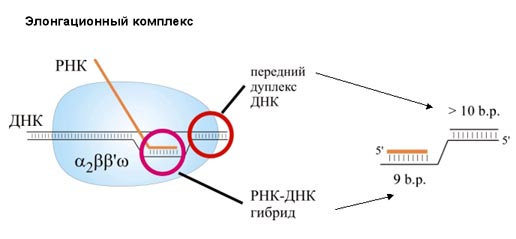
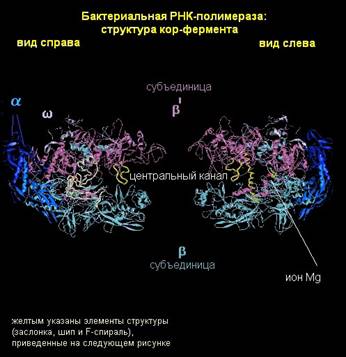
Разберем более подробно молекулярную структуру кор-фермента. Как было сказано выше, кор-фермент состоит из α- и β-субъединиц. Они соединены так, что образуют как бы «пасть» или «клешню». α-субъединицы находятся в основании этой «клешни», и выполняют структурную функцию. С ДНК и РНК они, по-видимому, не взаимодействуют. ω-субъединица – небольшой белок, который также выполняет структурную функцию. Основная часть работы приходится на долю β- и β΄-субъединиц. На рисунке β΄-субъединица показана наверху, а β-субъединица - внизу.
Внутри «пасти», которая называется главным каналом, находится активный центр фермента. Именно здесь происходит соединение нуклеотидов, образование новой связи при синтезе РНК. Главный канал в РНК-полимеразе – это то место, где во время элонгации находится ДНК. Еще в этой структуре сбоку есть так называемый вторичный канал, по которому подаются нуклеотиды для синтеза РНК.
Распределение зарядов на поверхности РНК-полимеразы обеспечивает ее функции. Распределение очень логично. Молекула нуклеиновой кислоты заряжена отрицательно. Поэтому полость главного канала, где должна удерживаться отрицательно заряженная ДНК, выложена положительными зарядами. Поверхность РНК-полимеразы выполнена отрицательно заряженными аминокислотами, чтобы ДНК к ней не прилипала.
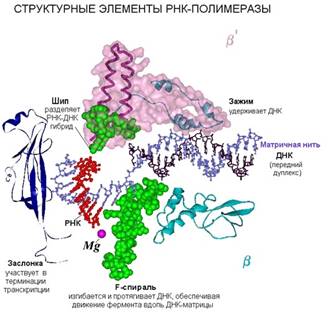
РНК-полимераза работает как молекулярная машина, и в ней есть различные детали, каждая из которых выполняет свою функцию. Например, нависающая над "пастью" часть β΄- субъединицы удерживает передний ДНК-дуплекс. Эта часть называется "заслонкой". После связывания с ДНК заслонка опускается, проходя путь в 30 ангстрем, и зажимает ДНК так, чтобы она не могла выпасть в процессе транскрипции.
внутри "пасти" находится активный центр РНК-полимеразы, то есть то место, где непосредственно происходит комплементарное взаимодействие поступившего по боковому каналу рибонуклеоиздтрифосфата с ДНК-матрицей. Если вновь прибывший нуклеотид комплементарен матрице, то он ферментативно пришивается к свободному 3' –концу РНК. По характеру реакция образования новой связи в РНК относится к реакциям нуклеофильного замещения. В ней участвуют два иона магния. Один ион постоянно находится в активном центре, а второй ион магния поступает с нуклеотидом и после образования новой связи между рибонуклеотидами уходит, затем поступает новый нуклеотид со своим новым ионом магния.
При выходе из РНК-полимеразы ДНК-РНК гибрид должен быть расплетен. В этом участвует структура, называемая "шип".
В транслокации, то есть перемещении РНК-полимеразы по нити ДНК, участвует α-спиральная структура, снизу вверх торчащая из β-субъединицы.
Как же узнали, какая часть фермента какую роль выполняет. Молекулярные биологи поступают следующим образом. Они удаляют часть белковой последовательности и смотрят, какая функция исчезла. Было показано, что если выбросить фрагмент зажима (когда его выбрасывали, еще не знали, что он держит ДНК), то ДНК держаться не будет. Такой же результат получается, если удалить ДНК переднего дуплекса. Оставшаяся часть - РНК-ДНК гибрид и задний дуплекс – оказываются слабо связанными с РНК-полимеразой.
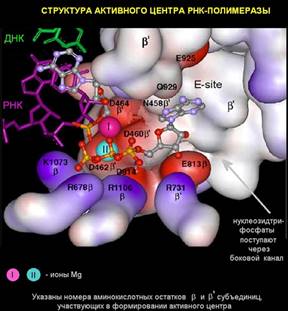
Известно, что магний координирует связь между фосфатами растущей молекулы ДНК и фосфатами вновь входящих нуклеотидов. При этом происходит последовательность реакций, называемых реакциями нуклеофильного замещения. Известно, каким образом меняются связи внутри этого комплекса. Новый нуклеотид приходит, будучи связанным с еще одним ионом магния. Новый нуклеотид таким образом взаимодействует с растущей цепью ДНК. В конце реакции, второй ион магния выводится из активного центра фермента.
РНК-полимераза является представителем молекулярных машин. Помимо того, что в начале синтеза ДНК опускается заслонка, меняется конформация других частей РНК-синтазы, в ней во время роста цепи РНК происходят циклические изменения, не такие сильные, как при начале синтеза цепи. В начале заслонка опускается на 30 Ǻ, а при каждом шаге фермента ДНК протягивается на один нуклеотид. В перемещении по ДНК участвует элемент РНК-полимеразы F-спираль (альфа-спиральная структуры, точащая из бета-субъединицы вверх в главный канал). F-спираль при этом изгибается, перемещается вместе с комплексом РНК-ДНК, освобождается от них и опять выпрямляется. Перемещается F-спираль за один шаг на 3,4 Ǻ. Именно такой шаг у РНК-полимеразы.
Изменение конформации различных частей РНК-полимеразы происходит за счет изменения потенциальной энергии, что связано с электростатическими и гидрофобными взаимодействиями. Можно провести следующую аналогию. Если взять поднос с горкой яблок, то после того, как мы этот поднос потрясем, яблоки будут рассыпаться ровным слоем по подносу. У них при этом изменится потенциальная энергия, связанная с действием силы тяжести. Если молекулу РНК-синтазы «потрясти» (а «трясет» ее, также как и все другие молекулы в клетке, броуновское движение), то она начнет принимать конформацию с более низкой потенциальной энергией. То есть, источником движения молекулярной машины является энергия теплового движения отдельных ее составляющих, а устройство машины таково, что это движение приводит к нужному результату. При этом молекулярная машина потребляет энергию, которая, в основном, идет на изменение состояния тех или иных связей.
Сейчас остановимся на инициации транскрипции. Как уже говорилось, инициация осуществляется с участием σ-субъединицей. Она взаимодействует со структурой ДНК, которая называется промотор. Она имеет у кишечной палочки такую структуру. За десять нуклеотидов до точки инициации находится ТАТА-бокс. Не обязательно стоит именно такая последовательность, но она является "идеальной" последовательностью для взаимодействия с σ-субъединицей, то есть такой, с которой транскрипция инициируется наиболее эффективно. Замена отдельных нуклеотидов в этой последовательности снижает эффективность инициации транскрипции. Еще примерно за 35 нуклеотидов до него находится структура, называемая «-35». Эту последовательность также распознает σ-субъединица. Эту структуру (сочетание последовательностей "–10" и "–35") назвали классическим промотором, т.к. она была описана первой. Но оказалось, что устройство промотора может быть и другим. Этот вариант включает в себя тот же ТАТА-бокс, но нет последовательности «-35», однако есть дополнительно два нуклеотида, и этого достаточно, чтобы σ-субъединица распознала промотор.
Эта структура называется расширенным промотором. σ-субъединица РНК-полимеразы садится на промотор в ДНК и разными частями белковой молекулы взаимодействует с частями промотора. Распознает его σ-субъединица через большую бороздку ДНК. После того, как σ-субъединица в составе кор-фермента связалась с промотором, ДНК на этом участке начинает плавиться (расплетаются нити ДНК). На прошлой лекции обсуждалось, что в паре А-Т связи между нуклеотидами разрываются легче, чем в паре Г-Ц, так как последняя содержит 3 водородных связи, а первая – две. Промотор содержит пары А-Т, поэтому плавится он достаточно легко. И затем начинается синтез РНК, растущая цепь РНК выталкивает σ-субъединицу и происходят еще другие изменения, которые вызывают диссоциацию σ-субъединицы от кор-фермента.
Теперь приведем пример, как изучают функции разных частей белка. Если небольшой кусочек белка отрезать и посмотреть, как изменились функции белка, то можно понять, какие были функции у отрезанного кусочка. В нашем случае сделали по-другому. Взяли две ДНК-полимеразы, одну взяли из кишечной палочки, а другую – из теплолюбивой бактерии (термофильной), которая растет при 800 С, (в лабораторных условиях их растят в колбе, которая находится в термостате в почти кипящей воде, в естественных условиях они живут в горячих источниках, есть такие, которые могут жить при 98оС), следовательно оптимум работы ее РНК-полимеразы и σ-субъединицы – 80оС, (на рисунке σ-субъединица термофильной бактерии показана красным, а кишечной палочки - желтым), а у кишечной палочки наиболее эффективная работа идет при температуре человеческого тела, (так как она живет в кишечнике). У ее σ-субъединицы всего четыре части, разрезали белок и сшивали эту σ-субъединицу с кусочком от σ-субъединицы термофильной бактерии. И потом разные кусочки от термофильной бактерии вставляли, заменяя ими разные фрагменты σ-субъединицы. Затем смотрели, активен ли полученный гибридный белок при 200 С или нет. Термофильная бактерия при такой температуре не работает, для нее это слишком холодно, а кишечная палочка активна. На рисунке видно, что при данной температуре работает только та комбинация, при которой у σ-субъединицы первая и вторая часть от кишечной палочки, а третья и четвертая от термофильной бактерии. Таким образом, делают вывод, что температуру работы σ-субъединицы определяют первая и вторая составные части.
На самом деле разрезают не белок, а ДНК, потом кусочки ДНК от разных бактерий сшивают вместе и затем вводят в бактерию, там при активизации этой части ДНК синтезируется гибридный белок. Эта технология относится к генной инженерии, она была разработана в 70-х годах.
Еще одной особенностью транскрипции является то, что кор-фермент бактериальной клетки один и тот же, а σ-субъединицы могут быть разными. У кишечной палочки всего 7 σ-субъединиц, они узнают разные промоторы. Зачем это нужно? Если клетке срочно нужно переключить синтез белков с одной группы генов на другую, она может использовать разные σ-субъединицы. Например, есть гены теплового шока, если кишечную палочку подогреть до состояния, когда жить ей станет очень тяжело, она включает аварийную систему сопротивления тепловому шоку, сопротивления тем разрушениям, которые произошли в клетке. В эту систему входит тот набор генов, который в норме работать не должен, перед этими генами свой особый промотор. И тогда другая σ-субъединица , не основная, синтезируется и активирует эти гены. То есть смена субъединицы – это смена программы работы генов. Это способ регуляции работы генов.
Трансляция
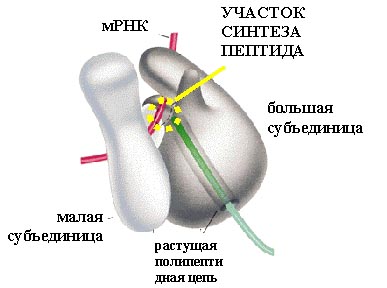
Перейдем к трансляции – синтезу белков. Она проводится рибосомами. Рибосома состоит из двух субчастиц: большой и малой.
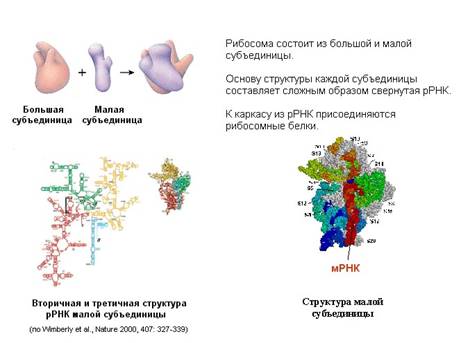
Каждая субчастица состоит из нескольких десятков белков, каждый из которых уже изучен, известно, каким образом каждый белок уложен в субчастицу. При исследовании белков используют метод электрофореза, то есть в электрическом поле в специальном геле или специальном носителе молекулы белков разъединяются в зависимости от их заряда и молекулярного веса, то есть под действием поля они начинают двигаться и могут отодвигаться друг от друга на разное расстояние. Другим методом разделения белков является хроматография, в результате этого метода на носителе получают пятнышки, каждый из которых соответствует отдельному белку.
Белки в рибосоме держатся на каркасе, состоящем из рибосомной РНК. Формирование рибосомы начинается с того, что рибосомная РНК сворачивается и на нее в определенном порядке начинают налипать белки. На рисунке представлена рибосомная РНК. В ней самокомплементарные участки нити РНК спариваются, образуя шпильки (вторичная структура), и затем РНК сворачивается (третичная структура РНК), образуя каркас субчастиц.
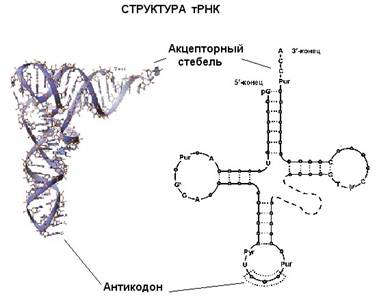
Еще один вид РНК, участвующей в синтезе белка, это транспортная РНК (тРНК). Молекулы тРНК относительно небольшие (по сравнению с рибосомногй или матричной РНК). Все тРНК имеют общую вторичную структуру. За счет спаривания комплементарных участков молекулы тРНК образуется три "стебля" с петлями на концах и один "стебель", образованный 5'- и 3'-концами молекулы тРНК (иногда образуется еще дополнительная пятая петля). Изображение этой структуры похоже на крест или клеверный лист. "Голова" на этом листе представлена антикодонной петлей, здесь находится антикодо – те три нуклеотида, которые комплементарно взаимодействуют с кодоном в мРНК. Противоположный антикодонной петле стебель, образованный концами молекулы, называется акцепторным стеблем – сюда присоединяется соответствующая аминокислота. Распознают подходящие друг другу тРНК и аминокислоты специальные ферменты, называемые аминоацил-тРНК синтетазами. Для каждой аминокислоты есть своя аминоацил-тРНК синтетаза.
В рибосоме находится матричная РНК (мРНК). С кодоном (тремя нуклеотидами) мРНК комплементарно связан антикодон транспортной РНК, на которой висит остаток аминокислоты. На рисунке видна такая структура (тРНК вместе с аминокислотой, которая называется аминоцил-тРНК).
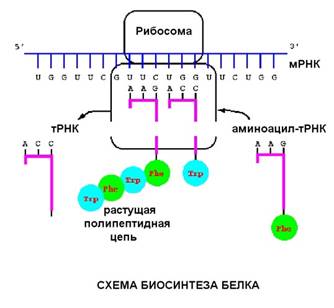
Процесс трансляции, также как и процесс транскрипции, связан с перемещением вдоль молекулы нуклеиновой кислоты, разница в том, что рибосома шагает на три нуклеотида, в то время как РНК-полимераза - на один.
Аминоцил т-РНК входит в рибосому, комплементарно связываясь с кодоном мРНК, затем происходит реакция при которой аминокислотные остатки связываются друг с другом, а т-РНК удаляется.
"Словарь" для перевода с языка нуклеотидов на язык аминокислот называется генетическим кодом. Аминокислот - 20, нуклеотидов – 4, число комбинаций из 4 по 2 = 16, а аминокислот 20, поэтому кодировка не двух, а трехбуквенная, каждая тройка называется кодоном. Каждая аминокислота кодируется тремя нуклеотидами в мРНК (которая, в свою очередь, кодируется ДНК).
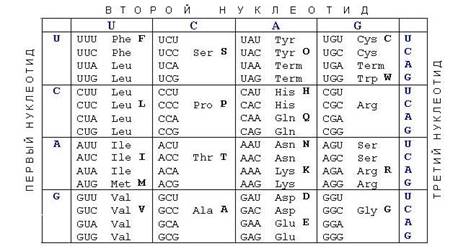
В таблице на рисунке боковые столбцы кодируют левую и правую букву кодона, верхняя строка – среднюю. Например кодон AUG кодирует аминокислоту метионин. Число комбинаций из 4 по 3 = 64, то есть некоторые аминокислоты кодируются несколькими кодонами. Три кодона не кодируют никакую аминокислоту, они называются терминирующими. Когда они попадаются в мРНК, рибосома прекращает свою работу и готовая полипептидная цепь выбрасывается наружу.
Таблица генетического кода была составлена в 60-х годах. Начало положили Ниренберг и Маттеию. Они пытались производить в пробирке эксперименты на клеточных экстрактах, к которым были добавлены искусственные матрицы РНК. В то время считалось, что кодоны, состоящие из одного нуклеотида (UUU или ААА) не кодируют аминокислоты. Ниренберг и Маттеи использовали полиU-РНК (то есть состоящую только из урацилов) в качестве контроля в своих опытах, но именно в этой пробирке прошла реакция. Стало ясно, что кодон UUU кодирует аминокислоту фенилаланин. Затем была составлена таблица генетического кода.
Генетический код универсален. Он один и тот же у всех микроорганизмов. Есть небольшие отличия в генетическом коде митохондрий.
Генетическим кодом называется таблица соответствия кодонов аминокислотам. Когда журналисты пишут о том, что недавно расшифрован генетический код человека – это грубая терминологическая ошибка. Генетический код человека расшифрован тогда же, когда и всех остальных живых существ – в 60-х годах XX века. Недавно расшифрован геном человека, то есть полная последовательность нуклеотидов всех молекул ДНК.
В лекции использованы изображения РНК-полимеразы, предоставленные Андреем Кульбачинским (Институт молекулярной генетики РАН).
Похожие рефераты:
Являются ли вирусы живыми организмами
рДНК-биотехнология. Способы биотрансформации клеток
Структура транскриптонов и транскрипция про- и эукариот
Литература - Другое (книга по генетике)
Репликация, сохранение и модификация генома
ДНК. Основы генетического материал
Продукты рекомбинации: характеристика и манипулирование
Структурно-функциональная организация генетического материала